В) фаза следовой реполяризации (потенциал) — Студопедия
В) фаза следовой реполяризации (потенциал)
Изменение проницаемости мембраны для Na+ продолжается недолго. Она начинает повышаться для K+ и снижается для Na+. Это соответствует фазе реполяризации. Нисходящая часть кривой соответствует следовому потенциалу и отражает восстановительные процессы наступающие после раздражения.
На кривой потенциала действия выделяют следующие фазы:
1.Локальный ответ (местная деполяризация), предшествующий развитию ПД.
2.Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше 0. Поэтому мембрана приобретает противоположный заряд – внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мсек.
3.Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мсек.
4.Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мсек.
5.Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу, МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мсек.
Амплитуда потенциала действия скелетных мышц в среднем 120-130 мВ, нейронов 80-90 мВ, гладкомышечных клеток 40-50 мВ. При возбуждении нейронов ПД возникает в начальном сегменте аксона – аксонном холмике.
Возникновение ПДобусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных зависит величина критического уровня деполяризации: чем она выше, тем ниже КУД и наоборот.
Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ), сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию.
Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой.
Следовая гиперполяризация связана с повышенной, после ПД, калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос, выносящий вошедшие в клетку во время ПД ионы натрия.
Изменяя проводимость быстрых натриевых и калиевых каналов можно влиять на генерацию ПД, а следовательно на возбуждение клеток. При полной блокаде натриевых каналов, например ядом рыбы тетродонта – тетродотоксином, клетка становится невозбудимой. Это используется в клинике. Такие местные анестетики, как новокаин, дикаин, лидокаин тормозят переход натриевых каналов нервных волокон в открытое состояние. Поэтому проведение нервных импульсов по чувствительным нервам прекращается, наступает обезболивание (анестезия) органа. При блокаде калиевых каналов затрудняется выход ионов калия из цитоплазмы на наружную поверхность мембраны, т.е. восстановление МП. Поэтому удлиняется фаза реполяризации. Этот эффект блокаторов калиевых каналов также используется в клинической практике. Например, один из них хинидин, удлиняя фазу реполяризации кардиомиоцитов, урежает сердечные сокращения и нормализует сердечный ритм.
Также следует отметить, что чем выше скорость распространения ПД по мембране клетки, ткани, тем выше ее проводимость.
http://studopedia.ru/19_315057_v-faza-sledovoy-repolyarizatsii-potentsial.html
Фазы реполяризации потенциала действия
Потенциалы действия, зарегистрированные в волокнах Пуркинье и в некоторых волокнах миокарда желудочков, имеют короткую, быструю фазу реполяризации (фаза 1), следующая непосредственно за фазой нарастания (см. рис. 3.1). Во время этой фазы мембранный потенциал временно возвращается почти к нулевому уровню, от которого начинается фаза плато потенциала действия, поэтому между этими двумя фазами на кривой иногда наблюдается четкий изгиб. Как было показано (в волокнах Пуркинье), быстрая реполяризация обусловлена транзиторным всплеском выходящего тока [30]. Во время нарастания потенциала действия этот выходящий ток активируется при деполяризации до уровня положительного потенциала, после чего он инактивируется как вследствие зависимого от времени процесса, так и в результате реполяризации. Хотя раньше считалось, что этот выходящий ток переносится преимущественно ионами хлора, в настоящее время более вероятным представляется его перенос главным образом ионами калия и лишь отчасти — ионами хлора [29].
Во время фазы плато потенциала действия, которая может длиться сотни миллисекунд, скорость реполяризации мембраны значительно ниже, так как суммарная величина выходящего мембранного тока невелика; входящие токи, сохраняющиеся в результате неполной инактивации натриевых и кальциевых каналов, приблизительно сбалансированы направленными наружу мембранными токами [30, 31]. По крайней мере один из них, вероятнее всего, является калиевым током, проходящим через ворота каналов, проводимость которых зависит от времени и потенциала. Активация их проводимости (только медленная) отмечается на уровне плато мембранного потенциала. Небольшой вклад в выходящий (реполяризующий) мембранный ток при этом уровне потенциала вносит и направленное внутрь движение ионов хлора, а также активность Na—К-насоса, генерирующего суммарный выходящий ток Na + [39]. По мере того как суммарный трансмембранный ток на уровне потенциала плато (т. е. алгебраическая сумма всех компонентов входящего и выходящего токов) становится более выходящим, мембранный потенциал все быстрее смещается в отрицательном направлении и начинается конечная быстрая фаза реполяризации потенциала действия. Эта конечная реполяризация, как и начальная фаза быстрой деполяризации, является регенеративной, но в отличие от фазы нарастания она, вероятно, включает изменения проводимости, зависящие главным образом от потенциала, а не от времени, и, следовательно, отражает время, затрачиваемое выходящим ионным током на обеспечение необходимой проводимости мембраны [34].
Спонтанная диастолическая деполяризация и автоматизм
Мембранный потенциал нормальных клеток рабочего миокарда предсердий и желудочков остается постоянным на уровне потенциала покоя в течение всей диастолы (см. рис. 3.1): если эти клетки не возбуждаются распространяющимся импульсом, то потенциал покоя в них поддерживается сколь угодно долго. В сердечных волокнах другого типа, например в специализированных волокнах предсердий или в волокнах Пуркинье проводящей системы желудочков, мембранный потенциал во время диастолы непостоянен и постепенно изменяется в сторону деполяризации. Если такое волокно не будет возбуждено распространяющимся импульсом раньше, чем мембранный потенциал достигнет порогового уровня, то в нем может возникнуть спонтанный потенциал действия (рис. 3.6). Изменение мембранного потенциала во время диастолы называется спонтанной диастолической деполяризацией, или фазой 4 деполяризации. Обусловливая возникновение потенциалов действия, этот механизм служит основой автоматизма. Автоматизм является нормальным свойством клеток синусового узла, мышечных волокон митрального и трикуспидального клапанов, некоторых участков предсердий, дистальной части АВ-узла, а также тканей системы Гиса — Пуркинье. В здоровом сердце частота возникновения импульсов вследствие автоматизма клеток синусового узла достаточно высока, что позволяет распространяющимся импульсам возбуждать другие потенциально автоматические клетки, прежде чем они спонтанно деполяризуются до порогового уровня. При этом потенциальная автоматическая активность других клеток обычно подавляется, хотя при целом ряде физиологических и патологических состояний она может проявляться (обсуждается ниже).

Рис. 3.6.Спонтанная диастолическая деполяризация и автоматизм волокон Пуркинье у собаки.
А — спонтанное возбуждение волокна Пуркинье при максимальном диастолическом потенциале —85 мВ. Диастолическая деполяризация является следствием уменьшения во времени тока ins, или пейсмекерного тока (см. в тексте). Б — автоматическая активность, возникающая при снижении мембранного потенциала; регистрация в волокне Пуркинье, перфузируемом безнатриевым раствором, но аналогичная активность наблюдается и в нормальном, содержащем ионы ^Vв + растворе Тироде. Фрагмент Б1: при деполяризации волокна (стрелка) от уровня потенциала покоя —60 до —45 мВ путем пропускания длительного импульса тока через микроэлектрод возникают три спонтанных потенциала действия. Фрагмент Б2: при большей амплитуде импульса мембранный потенциал снижается до —40 мВ, обусловливая поддерживающуюся ритмическую активность. Фрагмент Б3: усилившийся импульс тока снижает мембранный потенциал до —30 мВ, вследствие чего поддерживающаяся ритмическая активность возникает с большей частотой. Такая ритмическая активность, возникающая при менее негативных, чем —60 мВ, потенциалах, зависит, вероятно, от другого пейсмекерного тока, нежели активность, показания на фрагменте А.
Спонтанная диастолическая деполяризация является следствием постепенного изменения баланса между входящими и выходящими мембранными токами в пользу суммарного входящего (деполяризующего) тока. При исследовании пейсмекерного тока методом фиксации потенциала в волокнах Пуркинье [34—37] и клетках узла [38] была показана зависимость характеристик воротной системы как от потенциала, так и от времени. На основании данных первоначальных исследований уровня потенциала, при котором пейсмекерный ток реверсивно меняет свое направление, предполагалось, что выходящий пейсмекерный ток, переносимый ионами K + , постепенно отклоняется, позволяя тем самым направленному внутрь фоновому току деполяризовать клеточную мембрану [34—36]. Однако, согласно интерпретации результатов более поздних экспериментов, нормальным пейсмекерным током является входящий ток, переносимый преимущественно ионами Na + , который с течением времени возрастает, вызывая таким образом постепенную диастолическую деполяризацию [37, 38]. Когда деполяризация достигает уровня порогового потенциала, возникает импульс, после чего пейсмекерная проводимость при деполяризации мембраны инактивируется и сможет реактивироваться лишь после реполяризации потенциала действия. Ясно, что частота спонтанных возбуждений определяется временем, за которое диастолическая деполяризация изменяет мембранный потенциал до порогового уровня; следовательно, изменения порогового потенциала или скорости диастолической деполяризации, возникающие, например, в волокнах Пуркинье под действием адреналина, могут повлиять на частоту автоматической активности.
http://studfiles.net/preview/1209646/page:20/
Фаза реполяризации потенциала действия обусловлена диффузией ионов
Здравствуйте читатели моего проекта \»Биология для студентов\»! Подготовка к экзаменам, зачетам и госэкзаменам, а также рефераты и презентации занимают много времени, если готовится по учебникам. Есть три способа подготовки к экзамену: по учебнику, по лекциям и поиск в интернете. Готовиться по учебнику очень долго. Что касается лекций, не у всех есть хорошие лекции, так как не все преподаватели их нормально читают, и кроме того не все успевают их записывать. И остается третий вариант искать ответы на вопросы в интернете. Не для кого не секрет, что в настоящее время большинство студентов предпочитают именно этот вариант.

За пять лет учебы на факультете биотехнологии и биологии подготовка к сессии у меня занимала много времени. В Рунете не так много биологических сайтов. Конспекты по экономике, истории, социологии, политологии, математике найти очень просто. А ответы на вопросы по ботанике, зоологии, генетики, биофизике, биохимии гораздо сложнее. Наверное, потому что биология не самая распространенная специальность. К тому же биологические предметы не являются общеобразовательными в отличие, например, от экономики и истории, которые изучаются практически на любых специальностях. В Рунете я не нашел ни одного сайта на которым был бы представлен необходимый контент для подготовки к экзаменам, зачетам и госэкзаменам по биологическим дисциплинам. И я решил создать его.
Также я хотел бы вас попросить рассказать об этом сайте своим однокурсникам, друзьям и знакомым, которые являются студентами биологических специальностей. Это поможет развитию данного проекта.
http://vseobiology.ru/biofizika/1232-038-fazy-depolyarizatsii-repolyarizatsii-i-giperpolyarizatsii
Фазы деполяризации и реполяризации потенциала действия
В клетках синусового и атриовентрикулярного узлов скорость деполяризации в нулевую фазу намного ниже (1—20 В/с), чем в нормальных волокнах Пуркинье или клетках рабочего миокарда (см. рис. 3.9). Амплитуда потенциалов действия также весьма невелика (60—80 мВ); в некоторых клетках пик потенциала действия не превышает 0 мВ [3]. По сравнению с другими сердечными клетками более низкая скорость нарастания и меньшая амплитуда потенциала действия узловых клеток отражают значительно меньшую величину входящего тока перед нулевой фазой деполяризации в этих клетках. Имеющиеся в настоящее время данные однозначно свидетельствуют о том, что меньший по величине входящий ток в клетках синусового и атриовентрикулярного узлов течет не через быстрые натриевые каналы, а через медленные каналы и переносится ионами натрия и кальция [45—47]. Такие потенциалы действия с нарастанием, зависящим от медленного входящего тока, часто называют «медленными ответами» в отличие от более обычных «быстрых ответов», нарастание в которых зависит от быстрого натриевого тока [20]. Из-за столь малой величины суммарного входящего тока и медленной деполяризации в нулевую фазу скорость проведения медленных потенциалов действия через узлы всегда низка (0,01—0,1 м/с); именно такое медленное проведение в определенных условиях может обусловить возникновение нарушений ритма в тканях узлов. Как отмечалось ранее, медленные каналы для входящего тока имеют совершенно иные характеристики зависимости их воротного механизма от времени и потенциала по сравнению с быстрыми натриевыми каналами. Медленный входящий ток активируется и инактивируется значительно дольше, чем быстрый натриевый ток. Поэтому после нарастания потенциала действия в узлах медленный входящий ток инактивируется только медленно, способствуя деполяризации мембраны в течение всей фазы плато потенциала действия. Активация зависимого от времени и потенциала выходящего калиевого тока вместе с инактивацией медленного входящего тока, по-видимому, вызывает реполяризацию клеток узлов, как это было описано для окончания потенциала действия в других сердечных клетках.
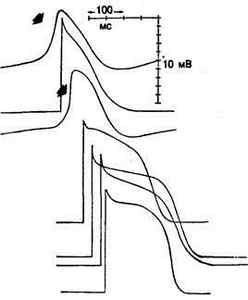
Рис. 3.9. Сравнение потенциалов действия синусового и АВ-узла (указаны стрелками) с потенциалами действия рабочего миокарда и волокон Пуркинье.
Запись потенциалов действия произведена в следующих областях сердца (начиная сверху): синусовый узел, предсердие, атриовентрикулярный узел, пучок Гиса, волокно Пуркинье в ложном сухожилии, терминальное волокно Пуркинье и рабочий миокард желудочков. Заметьте, что возрастание скорости и амплитуда потенциалов действия синусового и АВ-узла меньше аналогичных параметров в других клетках [3].
Проводимость медленных каналов для входящего тока восстанавливается после реполяризации мембраны также гораздо медленнее, чем проводимость быстрых натриевых каналов [20, 21]. В отличие от других сердечных клеток в узловых клетках при нанесении преждевременного стимула во время конечной фазы реполяризации не отмечается возникновения потенциала действия. В действительности достаточная инактивация проводимости медленных каналов для входящего тока может сохраняться даже после полной реполяризации в клетках, ставших абсолютно рефрактерными к стимуляции [48]. Реактивация происходит постепенно в течение всей диастолы; преждевременные импульсы, вызванные вскоре после полной реполяризации, имеют более медленное нарастание, меньшую амплитуду, чем нормальные импульсы, и распространяются медленнее. Преждевременные импульсы, вызванные позднее во время диастолы, имеют соответственно более быстрое нарастание, более высокую амплитуду и, следовательно, проводятся быстрее [49]. Такое поведение отражает длительность процесса реактивации медленных каналов. Связанный с этим продолжительный рефрактерный период узловой ткани, а также значительное замедление проведения через нее преждевременных импульсов могут быть важными факторами инициации некоторых нарушений сердечного ритма.
Автоматизм
Клетки синусового узла обычно являются автоматическими, так что каждый последующий потенциал действия возникает в результате спонтанной диастолической деполяризации; клетки АВ-узла также способны возбуждаться автоматически, особенно если они не связаны с окружающим миокардом предсердий [50]. Это означает, по-видимому, что электротоническое взаимодействие клеток узла и предсердий подавляется автоматически через предсердно-узловое проведение. Автоматическая активность клеток синусового узла не может быть связана с тем же пейсмекерным током, который был ранее описан в волокнах Пуркинье. Изменения состояния воротного механизма мембранной проводимости, ответственной за нормальный автоматизм волокон Пуркинье, происходят только в диапазоне мембранных потенциалов от —90 до —60 мВ [36]. Такие изменения проводимости вряд ли способны объяснить развитие спонтанной диастолической деполяризации в клетках синусового узла, поскольку максимальный диастолический потенциал этих клеток обычно менее отрицателен, чем —60 мВ. Однако имеющиеся данные говорят о том, что пейсмекерный ток в синусовом узле переносится, по крайней мере частично, ионами К + [38] и уменьшение этого выходящего тока при наличии постоянного фонового входящего тока приводит к постепенной деполяризации мембраны. Кроме того, важную роль играет, вероятно, входящий ток, который активируется при гиперполяризации и обозначается if [38].
http://infopedia.su/12x90c6.html
Фазы деполяризации потенциала действия
Электрический импульс, распространяющийся по сердцу и запускающий каждый цикл сокращений, называется потенциалом действия; он представляет собой волну кратковременной деполяризации, во время которой внутриклеточный потенциал поочередно в каждой клетке становится на короткое время положительным, а затем возвращается к своему исходному отрицательному уровню. Изменения нормального сердечного потенциала действия имеют характерное развитие во времени, которое для удобства подразделено на следующие фазы [3]: фаза 0 — начальная быстрая деполяризация мембраны; фаза 1 — быстрая, но неполная реполяризация; фаза 2 — «плато», или продолжительная деполяризация, характерная для потенциала действия сердечных клеток; фаза 3 — конечная быстрая реполяризация; фаза 4 — период диастолы.
При потенциале действия внутриклеточный потенциал становится положительным, так как возбужденная мембрана временно приобретает большую проницаемость для Na + (по сравнению с К + ),поэтому мембранный потенциал на какое-то время приближается по величине к потенциалу равновесия ионов натрия (ENa)- ЕNаможно определить, используя отношение Нернста; при внеклеточной и внутриклеточной концентрации Na + 150 и 10 мМ соответственно он составит:
Однако повышенная проницаемость для Na + сохраняется лишь непродолжительное время, так что мембранный потенциал не достигает ENaи после окончания потенциала действия возвращается к уровню покоя.
Указанные выше изменения проницаемости, вызывающие развитие фазы деполяризации потенциала действия, возникают вследствие открытия и закрытия особых мембранных каналов, или пор, через которые легко проходят ионы натрия. Как полагают, работа «ворот» регулирует открытие и закрытие отдельных каналов, которые могут существовать по меньшей мере в трех конформациях — «открытой», «закрытой» и «инактивированной». Одни ворота, соответствующие активационной переменной «m» в описании Ходжкина — Хаксли [13] ионных потоков натрия в мембране гигантского аксона кальмара, быстро перемещаются, открывая канал, когда мембрана внезапно деполяризуется под действием стимула. Другие ворота, соответствующие инактивационной переменной «h» в описании Ходжкина — Хаксли, при деполяризации движутся медленнее, и их функция заключается в закрытии канала (рис. 3.3). Как установившееся распределение ворот в пределах системы каналов, так и скорость их перехода из одного положения в другое зависят от уровня мембранного потенциала. Поэтому для описания мембранной проводимости Na + используются термины «зависимый от времени» и «потенциалозависимый».
Если мембрану в покое внезапно деполяризовать до уровня положительного потенциала (например, в эксперименте по фиксации потенциала), то активационные ворота быстро изменят свое положение, чтобы открыть натриевые каналы, а затем инактивационные ворота медленно их закроют (рис. 3.3). Слово «медленно» означает здесь, что на инактивацию уходит несколько миллисекунд, тогда как активация происходит в доли миллисекунды. Ворота остаются в указанных положениях до тех пор, пока мембранный потенциал снова не изменится, и для того чтобы все ворота вернулись к исходному состоянию покоя, мембрана должна быть полностью реполяризована до уровня высокого отрицательного потенциала. Если мембрана реполяризуется лишь до невысокого уровня отрицательного потенциала, то некоторые инактивационные ворота останутся закрытыми и максимальное число доступных натриевых каналов, способных открыться при последующей деполяризации, сократится [14]. (Электрическая активность сердечных клеток, в которых натриевые каналы полностью инактивированы, будет обсуждаться ниже.) Полная реполяризация мембраны в конце нормального потенциала действия обеспечивает возврат всех ворот к исходному состоянию и, следовательно, их готовность к следующему потенциалу действия.

Рис.3.3. Схематическое изображение мембранных каналов для входящих потоков ионов при потенциале покоя, а также при активации и инактивации.
Слева показана последовательность состояний канала при нормальном потенциале покоя —90 мВ. В покое инактивационные ворота как Na + -канала (h), так и медленного Ca 2+ /Na + -канала (f) открыты. Во время активации при возбуждении клетки т-ворота Na + -канала открываются и входящий поток ионовNa + деполяризует клетку, что приводит к нарастанию потенциала действия (график внизу). Затем h-ворота закрываются, инактивируя таким образом проводимостьNa + . При нарастании потенциала действия мембранный потенциал превышает более положительный порог потенциала медленных каналов; их активационные ворота (d) при этом открываются и ионы Ca 2+ и Na + поступают в клетку, вызывая развитие фазы плато потенциала действия. Ворота f, инактивирующие Ca 2+ /Na + -каналы, закрываются гораздо медленнее, чем ворота h, которые инактивируют Na-каналы. На центральном фрагменте показано поведение канала при снижении потенциала покоя до менее чем —60 мВ. Большинство инактивационных ворот Na-канала остается закрытым до тех пор, пока мембрана деполяризована; возникающий при стимуляции клетки входящий потокNa + слишком мал, чтобы вызвать развитие потенциала действия. Однако инактивационные ворота (f) медленных каналов при этом не закрываются и, как показано на фрагменте справа, при достаточном возбуждении клетки, позволяющем открыть медленные каналы и пропустить медленно входящие потоки ионов, возможно ответное медленное развитие потенциала действия.

Рис. 3.4.Пороговый потенциал при возбуждении сердечной клетки.
Слева — потенциал действия, возникающий на уровне потенциала покоя, составляющем —90 мВ; это происходит при возбуждении клетки поступающим импульсом или каким-либо подпороговым стимулом, быстро понижающим мембранный потенциал до значений, лежащих ниже порогового уровня в — 65 мВ. Справа — эффекты двух подпороговых и порогового стимулов. Подпороговые стимулы (а и б) не дают снижения мембранного потенциала до порогового уровня; следовательно, потенциал действия не возникает. Пороговый стимул (в) снижает мембранный потенциал точно до порогового уровня, на котором затем возникает потенциал действия [3].
Быстрая деполяризация в начале потенциала действия вызывается мощным входящим потоком ионов натрия, поступающих внутрь клетки (соответственно градиенту их электрохимического потенциала) через открытые натриевые каналы [6, 15]. Однако прежде всего натриевые каналы должны быть эффективно открыты, для чего требуется быстрая деполяризация достаточно большой площади мембраны до необходимого уровня, называемого пороговым потенциалом (рис. 3.4). В эксперименте этого можно достичь, пропуская через мембрану ток из внешнего источника и используя внеклеточный или внутриклеточный стимулирующий электрод. В естественных условиях той же цели служат локальные токи, протекающие через мембрану непосредственно перед распространяющимся потенциалом действия. При пороговом потенциале достаточное количество натриевых каналов оказывается открытым, что обеспечивает необходимую амплитуду входящего натриевого тока и, следовательно, дальнейшую деполяризацию мембраны; в свою очередь деполяризация вызывает открытие большего числа каналов, приводя к увеличению входящего потока ионов, так что процесс деполяризации становится регенеративным. Скорость регенеративной деполяризации (или нарастания потенциала действия) зависит от силы входящего натриевого тока, которая в свою очередь определяется такими факторами, как величина градиента электрохимического потенциала Na + и количество доступных (или неинактивированных) натриевых каналов. В волокнах Пуркинье максимальная скорость деполяризации при развитии потенциала действия, обозначаемая как dV/dtmaxили Vmax, достигает примерно 500 В/с, и если бы такая скорость поддерживалась в течение всей фазы деполяризации от —90 мВ до +30 мВ, то изменение потенциала на 120 мВ заняло бы около 0,25 мс. Максимальная скорость деполяризации волокон рабочего миокарда желудочков составляет примерно 200 В/с, а мышечных волокон предсердий—от 100 до 200 В/с [3]. (Фаза деполяризации потенциала действия в клетках синусового и атриовентрикулярного узлов существенно отличается от только что описанной и будет обсуждаться отдельно; см. ниже.)
Потенциалы действия с такой высокой скоростью нарастания (их часто называют «быстрыми ответами») быстро распространяются по сердцу. Скорость распространения потенциала действия (как и Vmax) в клетках с одинаковой пропускной способностью мембраны и характеристиками осевого сопротивления определяется главным образом амплитудой направленного внутрь тока, протекающего во время фазы нарастания потенциала действия. Это связано с тем, что локальные токи, проходящие через клетки непосредственно перед потенциалом действия, имеют большую величину при более быстром нарастании потенциала, поэтому мембранный потенциал в этих клетках раньше достигает порогового уровня, чем в случае токов меньшей величины (см. рис. 3.4). Конечно, эти локальные токи протекают через клеточную мембрану и сразу после прохождения распространяющегося потенциала действия, но они уже неспособны возбудить мембрану ввиду ее рефрактерности.

Рис.3.5. Нормальный потенциал действия и реакции, вызванные стимулами на разных стадиях реполяризации.
Амплитуда и возрастание скорости ответов, вызванных во время реполяризации, зависят от уровня мембранного потенциала, на котором они возникают. Самые ранние ответы (а и б) возникают на столь низком уровне, что оказываются слишком слабыми и неспособными к распространению (градуальные или местные ответы). Ответ «в» представляет собой наиболее ранний из распространяющихся потенциалов действия, но его распространение происходит медленно ввиду незначительного возрастания скорости, а также низкой амплитуды. Ответ «г» появляется точно перед полной реполяризацией, скорость его усиления и амплитуда выше, чем при ответе «в», так как он возникает при более высоком мембранном потенциале; однако скорость его распространения становится ниже нормальной. Ответ «д» отмечается после полной реполяризации, поэтому его амплитуда и скорость деполяризации имеют нормальные значения; следовательно, он быстро распространяется. ПП — потенциал покоя.
Продолжительный рефрактерный период после возбуждения сердечных клеток обусловлен большой длительностью потенциала действия и вольтажной зависимостью механизма ворот натриевых каналов. За фазой нарастания потенциала действия следует период продолжительностью от сотни до нескольких сотен миллисекунд, в течение которого регенеративный ответ на повторный стимул отсутствует (рис. 3.5). Это так называемый абсолютный, или эффективный, рефрактерный период; обычно он охватывает плато (фаза 2) потенциала действия. Как описано выше, натриевые каналы инактивируются и остаются закрытыми во время такой поддерживающейся деполяризации. В ходе реполяризации потенциала действия (фаза 3) инактивация постепенно устраняется, так что доля каналов, способных снова активироваться, постоянно возрастает. Следовательно, с помощью стимула в начале реполяризации можно вызвать лишь небольшой входящий поток ионов натрия, однако по мере продолжения реполяризации потенциала действия такие потоки будут увеличиваться. Если некоторые из натриевых каналов остаются невозбудимыми, то вызванный входящий поток Na + может привести к регенеративной деполяризации и, следовательно, к возникновению потенциала действия. Однако скорость деполяризации, а значит, и скорость распространения потенциалов действия значительно снижены (см. рис. 3.5) и нормализуются только после полной реполяризации [17, 18]. Время, в течение которого повторный стимул способен вызвать такие «градуальные» потенциалы действия, называется относительным рефрактерным периодом. Вольтажная зависимость устранения инактивации изучалась Weidmann, установившим, что скорость повышения потенциала действия и возможный уровень, при котором этот потенциал вызывается, находятся в S-образной зависимости, известной также как кривая реактивности мембраны.
Низкая скорость нарастания потенциалов действия, вызванных в течение относительного рефрактерного периода, обусловливает их медленное распространение; такие потенциалы действия могут послужить причиной некоторых нарушений проведения, например задержки, затухания и блокирования, и даже способны вызвать циркуляцию возбуждения. Данные явления обсуждаются ниже в этой главе.
В нормальных сердечных клетках за входящим натриевым током, ответственным за быстрое нарастание потенциала действия, следует второй входящий ток, меньшей величины и более медленный, чем натриевый ток, который, по-видимому, переносится в основном ионами кальция [19, 20]. Этот ток обычно относят к «медленному входящему току» (хотя он является таковым только в сравнении с быстрым натриевым током; другие важные изменения, например наблюдаемые во время реполяризации, вероятно, замедляются); он протекает через каналы, которые в соответствии с характеристиками их проводимости, зависящей от времени и вольтажа, были названы «медленными каналами» (см. рис. 3.3) [21]. Порог активации этой проводимости (т. е. когда начинают открываться активационные ворота — d) лежит между —30 и —40 мВ (сравните: от —60 до —70 мВ для натриевой проводимости) [20]. Регенеративная деполяризация, обусловленная быстрым натриевым током, обычно активирует проводимость медленного входящего тока, поэтому в более поздний период нарастания потенциала действия ток течет по каналам обоих типов. Однако ток Са 2+ гораздо меньше максимального быстрого тока Na + , поэтому его вклад в потенциал действия весьма невелик до тех пор, пока быстрый ток Na + не станет достаточно инактивированным (т. е. после начального быстрого нарастания потенциала). Поскольку медленный входящий ток может инактивироваться лишь очень медленно, он вносит свой вклад в основном в фазу плато потенциала действия. Так, уровень плато смещается в сторону деполяризации, когда градиент электрохимического потенциала для Са 2+ увеличивается при повышении концентрации [Са 2+ ]0; снижение [Са 2+ ]0вызывает смещение уровня плато в противоположную сторону [22, 23]. Однако в некоторых случаях может отмечаться вклад кальциевого тока в фазу нарастания потенциала действия. Например, на кривой нарастания потенциала действия в миокардиальных волокнах желудочка лягушки иногда наблюдается изгиб около 0 мВ, в точке, где первоначальная быстрая деполяризация уступает место более медленной деполяризации, которая продолжается до пика овершута потенциала действия. Как было показано, скорость более медленной деполяризации и величина овершута возрастают с повышением [Са 2+ ]0[23, 24].
Кроме различной зависимости от мембранного потенциала и времени, эти два типа проводимости различаются и по своим фармакологическим характеристикам. Так, ток через быстрые каналы для Na + снижается под действием тетродотоксина (ТТХ) [25], тогда как медленный ток Са 2+ не поддается влиянию ТТХ [20], но усиливается под действием катехоламинов [26] и угнетается ионами марганца [27], а также некоторыми препаратами, такими как верапамил и D-600 [28]. Представляется весьма вероятным (по крайней мере в сердце лягушки), что большая часть кальция, необходимого для активации белков, способствующих каждому сокращению сердца, попадает в клетку во время потенциала действия через медленный канал для входящего тока. У млекопитающих доступным дополнительным источником Са 2+ для сердечных клеток служат его запасы в саркоплазматическом ретикулуме.
http://studfiles.net/preview/1209646/page:19/
Фазы реполяризации потенциала действия
Потенциалы действия, зарегистрированные в волокнах Пуркинье и в некоторых волокнах миокарда желудочков, имеют короткую, быструю фазу реполяризации (фаза 1), следующая непосредственно за фазой нарастания (см. рис. 3.1). Во время этой фазы мембранный потенциал временно возвращается почти к нулевому уровню, от которого начинается фаза плато потенциала действия, поэтому между этими двумя фазами на кривой иногда наблюдается четкий изгиб. Как было показано (в волокнах Пуркинье), быстрая реполяризация обусловлена транзиторным всплеском выходящего тока [30]. Во время нарастания потенциала действия этот выходящий ток активируется при
деполяризации до уровня положительного потенциала, после чего он инактивируется как вследствие зависимого от времени процесса, так и в результате реполяризации. Хотя раньше считалось, что этот выходящий ток переносится преимущественно ионами хлора, в настоящее время более вероятным представляется его перенос главным образом ионами калия и лишь отчасти — ионами хлора [29].
Во время фазы плато потенциала действия, которая может длиться сотни миллисекунд, скорость реполяризации мембраны значительно ниже, так как суммарная величина выходящего мембранного тока невелика; входящие токи, сохраняющиеся в результате неполной инактивации натриевых и кальциевых каналов, приблизительно сбалансированы направленными наружу мембранными токами [30, 31]. По крайней мере один из них, вероятнее всего, является калиевым током, проходящим через ворота каналов, проводимость которых зависит от времени и потенциала. Активация их проводимости (только медленная) отмечается на уровне плато мембранного потенциала. Небольшой вклад в выходящий (реполяризующий) мембранный ток при этом уровне потенциала вносит и направленное внутрь движение ионов хлора, а также активность Na—К-насоса, генерирующего суммарный выходящий ток Na + [39]. По мере того как суммарный трансмембранный ток на уровне потенциала плато (т. е. алгебраическая сумма всех компонентов входящего и выходящего токов) становится более выходящим, мембранный потенциал все быстрее смещается в отрицательном направлении и начинается конечная быстрая фаза реполяризации потенциала действия. Эта конечная реполяризация, как и начальная фаза быстрой деполяризации, является регенеративной, но в отличие от фазы нарастания она, вероятно, включает изменения проводимости, зависящие главным образом от потенциала, а не от времени, и, следовательно, отражает время, затрачиваемое выходящим ионным током на обеспечение необходимой проводимости мембраны [34].
Спонтанная диастолическая деполяризация и автоматизм
Мембранный потенциал нормальных клеток рабочего миокарда предсердий и желудочков остается постоянным на уровне потенциала покоя в течение всей диастолы (см. рис. 3.1): если эти клетки не возбуждаются распространяющимся импульсом, то потенциал покоя в них поддерживается сколь угодно долго. В сердечных волокнах другого типа, например в специализированных волокнах предсердий или в волокнах Пуркинье проводящей системы желудочков, мембранный потенциал во время диастолы непостоянен и постепенно изменяется в сторону деполяризации. Если такое волокно не будет возбуждено распространяющимся импульсом раньше, чем мембранный потенциал достигнет порогового уровня, то в нем может возникнуть спонтанный потенциал действия (рис. 3.6). Изменение мембранного потенциала во время диастолы называется спонтанной диастолической деполяризацией, или фазой 4 деполяризации. Обусловливая возникновение потенциалов действия, этот механизм служит основой автоматизма. Автоматизм является нормальным свойством клеток синусового узла, мышечных волокон митрального и трикуспидального клапанов, некоторых участков предсердий, дистальной части АВ-узла, а также тканей системы Гиса — Пуркинье. В здоровом сердце частота возникновения импульсов вследствие автоматизма клеток синусового узла достаточно высока, что позволяет распространяющимся импульсам возбуждать другие потенциально автоматические клетки, прежде чем они спонтанно деполяризуются до порогового уровня. При этом потенциальная автоматическая активность других клеток обычно подавляется, хотя при целом ряде физиологических и патологических состояний она может проявляться (обсуждается ниже).
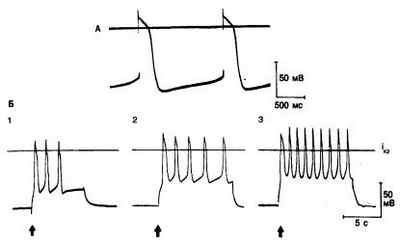
Рис. 3.6. Спонтанная диастолическая деполяризация и автоматизм волокон Пуркинье у собаки.
А — спонтанное возбуждение волокна Пуркинье при максимальном диастолическом потенциале —85 мВ. Диастолическая деполяризация является следствием уменьшения во времени тока ins, или пейсмекерного тока (см. в тексте). Б — автоматическая активность, возникающая при снижении мембранного потенциала;
р егистрация в волокне Пуркинье, перфузируемом безнатриевым раствором, но аналогичная активность наблюдается и в нормальном, содержащем ионы Na + растворе Тироде. Фрагмент Б1: при деполяризации волокна (стрелка) от уровня потенциала покоя —60 до —45 мВ путем пропускания длительного импульса тока через микроэлектрод возникают три спонтанных потенциала действия. Фрагмент Б2: при большей амплитуде импульса мембранный потенциал снижается до —40 мВ, обусловливая поддерживающуюся ритмическую активность. Фрагмент Б3: усилившийся импульс тока снижает мембранный потенциал до —30 мВ, вследствие чего поддерживающаяся ритмическая активность возникает с большей частотой. Такая ритмическая активность, возникающая при менее негативных, чем —60 мВ, потенциалах, зависит, вероятно, от другого пейсмекерного тока, нежели активность, показания на фрагменте А.
Спонтанная диастолическая деполяризация является следствием постепенного изменения баланса между входящими и выходящими мембранными токами в пользу суммарного входящего (деполяризующего) тока. При исследовании пейсмекерного тока методом фиксации потенциала в волокнах Пуркинье [34—37] и клетках узла [38] была показана зависимость характеристик воротной системы как от потенциала, так и от времени. На основании данных первоначальных исследований уровня потенциала, при котором пейсмекерный ток реверсивно меняет свое направление, предполагалось, что выходящий пейсмекерный ток, переносимый ионами K + , постепенно уменьшается, позволяя тем самым направленному внутрь фоновому току деполяризовать клеточную мембрану [34—36]. Однако, согласно интерпретации результатов более поздних экспериментов, нормальным пейсмекерным током является входящий ток, переносимый преимущественно ионами Na + , который с течением времени возрастает, вызывая таким образом постепенную диастолическую деполяризацию [37, 38]. Когда деполяризация достигает уровня порогового потенциала, возникает импульс, после чего пейсмекерная проводимость при деполяризации мембраны инактивируется и сможет реактивироваться лишь после реполяризации потенциала действия. Ясно, что частота спонтанных возбуждений определяется временем, за которое диастолическая деполяризация изменяет мембранный потенциал до порогового уровня; следовательно, изменения порогового потенциала или скорости диастолической деполяризации, возникающие, например, в волокнах Пуркинье под действием адреналина, могут повлиять на частоту автоматической активности.
Задержанная постдеполяризация и триггерная поддерживающаяся ритмическая активность
Наряду с автоматизмом существует и другой механизм, способный обеспечить ритмическую генерацию импульсов в нормальных сердечных клетках. Механизм инициации возбуждения зависит от задержанной постдеполяризации, поэтому
ритмически возникающие с его помощью спонтанные импульсы называют триггерными потенциалами действия [20, 39]. Как отмечалось выше, автоматическая активность характеризуется спонтанной генерацией каждого импульса. Следовательно, если автоматическая клетка не возбуждается распространяющимся импульсом, она не остается в покое, а подвергается спонтанной диастолической деполяризации до тех пор, пока не возникнет потенциал действия. Это согласуется с использованием прилагательного «автоматический», смысл которого можно расшифровать как «обладающий способностью к самостоятельному движению». И наоборот, если обладающее триггерной активностью волокно не возбуждается распространяющимся импульсом, то оно остается молчащим. Поскольку триггерный импульс — это импульс, возникающий после (и в результате) другого импульса, триггерная активность не может иметь место до тех пор, пока волокно не будет возбуждено по крайней мере одним распространяющимся импульсом. Триггерная активность является одной из форм ритмической активности, при которой каждый импульс возникает в результате предшествующего импульса, за исключением, разумеется, первого (побуждающего) потенциала действия, который должен быть вызван стимулом.
Рис. 3.7. Постдеполяризация и триггерная активность в предсердном волокне коронарного синуса у
Фрагмент А: однократная стимуляция волокна вызывает развитие одного потенциала действия, за которым следует постгиперполяризация (жирная стрелка), а затем—задержанная постдеполяризация (светлая стрелка). Фрагмент Б: запись с другой клетки; первый потенциал действия (слева) вызывается внешним стимулом, но следующая за ним задержанная постдеполяризация (черная стрелка) достигает порогового потенциала и вызывает первый спонтанный потенциал действия, за которым следуют другие спонтанные потенциалы действия; спонтанные импульсы являются триггерными импульсами, поэтому они представляют так называемую триггерную активность.
Триггерные импульсы обусловлены задержанной постдеполяризацией, амплитуда которой достаточно велика для доведения мембранного потенциала до порогового уровня. Задержанная постдеполяризация — это транзиторная деполяризация, наблюдающаяся после окончания потенциала действия, но возникающая благодаря именно этому потенциалу. В норме задержанная постдеполяризация была зарегистрирована в предсердных клетках митрального клапана [40], в клетках коронарного синуса [41] ив предсердных волокнах гребенчатых мышц [42]. Как показывает рис. 3.7, задержанной постдеполяризации часто предшествует постгиперполяризация: следующий за потенциалом действия мембранный потенциал на короткое время становится более отрицательным, чем непосредственно перед началом потенциала действия. При затухании этой постгиперполяризации мембранный потенциал временно становится более положительным, чем непосредственно перед началом потенциала действия. Кратковременность изменений такой постдеполяризации четко отличает ее от нормальной спонтанной диастолической (пейсмекерной) деполяризации, при которой мембранный потенциал монотонно изменяется до тех пор, пока не возникнет следующий потенциал действия.
Задержанная постдеполяризация обычно бывает подпороговой, но в определенных условиях она может превысить пороговый потенциал; если это происходит, вследствие постдеполяризации возникает спонтанный потенциал действия. В упомянутых выше предсердных волокнах катехоламины увеличивают амплитуду постдеполяризации, в результате чего достигается уровень порогового потенциала [40, 41]. Амплитуда подпороговой постдеполяризации весьма чувствительна и к частоте возникновения потенциала действия [39, 42]. Повышение частоты стимуляции увеличивает амплитуду постдеполяризации (рис. 3.8), и, наоборот, уменьшение ее
частоты приводит к снижению амплитуды. Кроме того, если преждевременный потенциал действия при стимуляции возникает с постоянной частотой, то следующая за ним постдеполяризация имеет большую амплитуду, чем та, которая отмечается после регулярного потенциала действия. Более того, чем раньше во время основного цикла возникает преждевременный потенциал действия, тем больше амплитуда преждевременной постдеполяризации. При достаточно высокой частоте постоянной стимуляции или после достаточно раннего преждевременного стимула постдеполяризация может достигнуть порога и вызвать нестимулированные потенциалы действия. Первый спонтанный импульс отмечается после более короткого интервала по сравнению с длительностью основного цикла, так как постдеполяризация, вследствие которой он возник, начинается вскоре после реполяризации предшествующего потенциала действия. Следовательно, спонтанный импульс вызывает еще одну постдеполяризацию, которая также достигает порогового уровня, обусловливая появление второго спонтанного импульса (см. рис. 3.8). Этот последний импульс вызывает следующую постдеполяризацию, которая инициирует третий спонтанный импульс, и так на протяжении всего времени триггерной активности. Триггерная активность может спонтанно прекратиться, и, если это происходит, за последним нестимулированным импульсом обычно следует одна или несколько подпороговых постдеполяризаций.
Рис. 3.8. Индукция триггерной активности в предсердном волокне митрального клапана у обезьяны.
На каждом фрагменте показаны лишь нижняя часть потенциалов действия. Горизонтальные линии на фрагментах I и II проведены на уровне—30 мВ, а на фрагменте III — на уровне — 20 мВ. фрагмент IA и 1Б: триггерная активность, возникшая в результате сокращения длительности основного стимуляционного цикла. IA: продолжительность цикла стимуляции составляет 3400 мс; и за каждым потенциалом действия следует подпороговая задержанная постдеполяризация. В начале фрагмента IБ длительность цикла стимуляции сокращена до 1700 мс; заметно постепенное повышение амплитуды постдеполяризации, следующей за каждым из первых 4 вызванных стимуляцией потенциалов действия. За последним вызванным потенциалом действия следует спонтанный потенциал действия, а затем поддерживающаяся ритмическая активность, частота которой выше, чем при стимуляции. IIА и IIБ: возникновение ритмической активности вследствие единственного вызванного импульса. IIА: после периода покоя за единственным вызванным потенциалом действия (стрелка) следует подпороговая постдеполяризация. IIБ: в несколько иных условиях — после одиночного вызванного потенциала действия (стрелка) отмечается поддерживающаяся ритмическая активность. IIIA и IIIБ:
в озникновение триггерной активности вследствие преждевременной стимуляции. IIIA: преждевременный импульс (стрелка) вызывается во время фазы реполяризации постдеполяризации, и амплитуда последующей постдеполяризации возрастает. IIIБ: за преждевременным импульсом (большая стрелка) следует постдеполяризация, которая достигает порога (маленькая стрелка) и приводит к появлению ряда триггерных импульсов [40].
http://studfiles.net/preview/4333815/page:16/
1 Биоэлектрические явления в возбудимых тканях Потенциал действия распространяется по клеточной мембране
1.2.Биоэлектрические явления в возбудимых тканях
1.Потенциал действия распространяется по клеточной мембране:
с затуханием, декрементом
+без затухания, без декремента
+без уменьшения амплитуды
Под потенциалом действия понимают:
+быстрое колебание потенциала мембраны с ее перезарядкой
быстрое колебание потенциала без перезарядки мембраны
правильного ответа нет
Увеличение абсолютной величины мембранного потенциала покоя называют:
Разность потенциалов между цитоплазмой и окружающим клетку раствором называют:
В фазу быстрой деполяризации потенциала действия проницаемость мембраны увеличена для ионов:
Как называют период полного отсутствия возбудимости в фазу быстрой деполяризации?
Основным признаком возбуждения является:
Локальный ответ возникает на:
+очень слабый раздражитель (допороговой величины)
раздражитель пороговой величины
Локальный ответ возникает, когда:
+деполяризация мембраны еще не достигла КУДа (критического уровня деполяризации)
деполяризация мембраны достигает КУДа (критического уровня деполяризации)
правильного ответа нет
Отличие потенциала действия от локального ответа заключается в том, что:
возникший процесс остается там, где он возник
+возникший процесс распространяется на соседние участки
нет правильного ответа
Электрические явления на уровне ткани в состоянии возбуждения:
потенциал покоя — ПП
потенциал действия — ПД
токи покоя (повреждения) – ТП
токи градиента основного обмена – ТГОО
+токи действия — ТД
Какой показатель не относится к фазам потенциала действия?
реверсия или овершут
Во время фазы «деполяризации» потенциала действия (ПД) происходит :
+уменьшение абсолютной величины мембранного потенциала
возвращение абсолютной величины мембранного потенциала (МП) к исходному уровню
увеличение абсолютной величины МП по сравнению с исходным уровнем
Во время фазы «реполяризации» потенциала действия (ПД) происходит :
уменьшение абсолютной величины мембранного потенциала
+возвращение абсолютной величины мембранного потенциала (МП) к исходному уровню
увеличение абсолютной величины МП по сравнению с исходным уровнем
Во время фазы «гиперполяризации» потенциала действия (ПД) происходит:
уменьшение абсолютной величины мембранного потенциала
возвращение абсолютной величины мембранного потенциала (МП) к исходному уровню
+увеличение абсолютной величины МП по сравнению с исходным уровнем
Во время фазы «быстрой деполяризации» потенциала действия (ПД) клеточной мембраны происходит :
открытие лишь части натриевых каналов клеточной мембраны
+открытие большинства натриевых каналов клеточной мембраны
правильного ответа нет
Массивный вход ионов натрия в фазу «деполяризации» потенциала действия (ПД) происходит:
+по концентрационному и электрохимическому градиенту
только по концентрационному градиенту
только по электрохимическому градиенту
Фаза потенциала действия, во время которой цитоплазма приобретает положительный заряд по отношению к наружному раствору, называется:
В какую фазу потенциала действия активируются и открываются калиевые каналы мембраны?
В процессе генерации потенциала действия (ПД) абсолютно рефрактерный период возбудимости наблюдается:
+во время быстрой деполяризации
во время реполяризации
во время следовой деполяризации
во время следовой гиперполяризации
В абсолютно рефрактерный период происходит:
+быстрая активация большинства натриевых каналов
последующая инактивация большинства натриевых каналов
значительное реполяризационное действие выходящих калиевых токов
Рефрактерный период возбудимости это:
сохранение возбудимости на определенном уровне
+снижение возбудимости вплоть до нуля
Инактивация натриевой системы в процессе генерации потенциала действия приводит клетку в состояние:
Нисходящая фаза потенциала действия связана с большей проницаемостью мембраны для ионов:
Концентрация каких ионов внутри клетки в покое больше:
правильного ответа нет
Концентрация каких ионов снаружи клетки в покое больше:
правильного ответа нет
Уменьшение абсолютной величины мембранного потенциала покоя при действии раздражителя пороговой величины называется:
В цитоплазме нервных и мышечных клеток концентрация каких ионов по сравнению с наружным раствором выше:
В фазу быстрой деполяризации потенциала действия проницаемость мембраны мышечного волокна увеличивается для ионов:
Фаза деполяризации потенциала действия (ПД) возникает в результате:
+открытия в мембране натриевых ионных каналов
закрытия в мембране натриевых ионных каналов
открытия калиевых ионных каналов
Стимулом для генерации потенциала действия (ПД) является:
Уровень деполяризации мембраны, при котором возникает потенциал действия (ПД) называется?
Какой фазе возбудимости соответствует фаза «деполяризации» потенциала действия нервной клетки?
правильного ответа нет
Какой фазе возбудимости соответствует фаза «реполяризации» потенциала действия нервной клетки?
правильного ответа нет
Вид биоэлектрических явлений на уровне клетки в покое:
Вид биоэлектрических явлений на уровне клетки при возбуждении:
токи покоя (повреждения)
токи градиента основного обмена
Где содержание ионов калия больше в состоянии покоя?
одинаково с обеих сторон клеточной мембраны
Где содержание натрия больше в состоянии покоя клетки?
одинаково с обеих сторон клеточной мембраны
Биоэлектрические явления, регистрируемые на уровне клетки:
+потенциал покоя — ПП
+потенциал действия — ПД
токи покоя (повреждения) — ТП
токи градиента основного обмена — ТГОО
токи действия — ТД
Биоэлектрические явления, регистрируемые на уровне ткани:
потенциал покоя — ПП
потенциал действия — ПД
+токи покоя (повреждения) — ТП
+токи градиента основного обмена — ТГОО
+токи действия — ТД
+постоянный выходящий калиевый ток в покое
входящий калиевый ток при возбуждении
правильного ответа нет
Изменение возбудимости клеток или тканей при действии постоянного тока называется:
правильного ответа нет
Положения полярного закона действия постоянного тока на ткань сформулированы:
Потеря возбудимости ткани при действии сильного постоянного тока вследствие выраженной гиперполяризации:
Какой из физиологических показателей дает более полную характеристику процесса возбуждения в возбудимых тканях в единицу времени?
правильного ответа нет
Наименьшую продолжительность времени, в течение которого должен действовать стимул в 2 реобазы, чтобы вызвать ответ, называют:
Закон, согласно которому возбудимая структура на пороговые и сверхпороговые раздражения отвечает максимально возможным ответом, называют:
Мерой лабильности возбудимых тканей является:
+количество потенциалов, генерируемых в единицу времени
амплитуда потенциалов, генерируемых возбудимой клеткой в единицу времени
правильного ответа нет
Время, в течение которого должен действовать раздражитель в одну реобазу, чтобы вызвать возбуждение, называют:
Минимальную силу раздражителя, который должен действовать в течение длительного, но определенного времени, чтобы вызвать возбуждение, называют:
Ткань обладает наименьшей возбудимостью, если разность между критическим уровнем деполяризации (КУД) и мембранным потенциалом равна:
Ткань обладает наибольшей возбудимостью, если разность между критическим уровнем деполяризации (КУД) и мембранным потенциалом равна:
К мерам возбудимости относятся:
+все ответы правильные
Наименее точная мера возбудимости:
Наиболее точная мера возбудимости:
Какая мера возбудимости используется в медицинской практике?
Как изменится амплитуда потенциала действия при его распространении в нервных волокнах?
+никак не изменится
Какой вид ионного транспорта обеспечивает фазу быстрой деполяризации потенциала действия (ПД)?
пассивный и активный
Что показывает прибор, измеряющий потенциал покоя (ПП) нервной клетки, если и микроэлектрод, и макроэлектрод находятся на наружной поверхности мембраны:
величину, равную ПП
Как изменится форма потенциала действия при замедлении процесса инактивации натриевых каналов?
укоротится фаза деполяризации
удлинится фаза деполяризации
укоротится фаза реполяризации
+удлинится фаза реполяризации
Как скажется действие препарата, способствующего дополнительному открытию калиевых каналов на продолжительности следовой гиперполяризации?
Если бы при раздражении нерва активация натриевых и калиевых каналов происходила не последовательно, а одновременно, к чему бы это привело?
+деполяризация бы не возникла
правильного ответа нет
Чем объяснить, что раздражители разной природы (механо-, хемо-, электро-) вызывают один и тот же эффект – возбуждение?
все они вызывают гиперполяризацию клеточной мембраны
+все они вызывают деполяризацию клеточной мембраны
В какую фазу потенциала действия (ПД) восстанавливается выходящий ток калия, инактивируется входящий ток натрия и активно работает калиево-натриевый насос?
Проводимость мембраны для ионов натрия начинает быстро снижаться:
в начале деполяризации мембраны
+после пика потенциала действия
в следовую деполяризацию
правильного ответа нет
Скорость и степень активации натриевых каналов:
+зависит от силы раздражителя
Деполяризация мембраны клетки:
+увеличивает число открытых потенциал зависимых натриевых каналов
уменьшает число открытых потенциал зависимых натриевых каналов
не оказывает влияния на число открытых потенциал зависимых натриевых каналов
Активация натриевой системы в процессе генерации потенциала действия (ПД) приводит клетку в состояние:
Достижение критического уровня деполяризации (КУД) мембраны :
+соответствует порогу активации потенциал зависимых натриевых каналов
соответствует порогу инактивации потенциал зависимых натриевых каналов
правильного ответа нет
потенциал зависимые натриевые каналы пока не инактивируются
+потенциал зависимые натриевые каналы уже инактивируются
правильного ответа нет
Какое раздражение способно вызвать потенциал действия в абсолютно рефрактерный период возбудимости?
+правильного ответа нет
Значительным реполяризационным действием выходящего калиевого тока и снижением инактивации натриевых каналов обусловлен:
абсолютно рефрактерный период
+относительно рефрактерный период
правильного ответа нет
Если искусственно снизить концентрацию ионов К+ внутри нервной клетки, величина потенциала покоя (ПП):
Потенциал покоя (ПП) обусловлен:
входом ионов калия (K)
входом ионов натрия (Na)
+выходом ионов калия (K)
выходом ионов натрия (Na)
Потенциал действия (ПД) обусловлен:
входом ионов калия (K)
+входом ионов натрия (Na)
выходом ионов калия (K)
выходом ионов натрия (Na)
Как изменится потенциал покоя (ПП), если уменьшить внутриклеточную концентрацию ионов K+?
Как изменится мембранный потенциал (МП), если заблокировать работу Na+ — К+ — зависимой АТФ-азы?
+станет равным нулю
Уменьшение величины мембранного потенциала покоя (ПП) при действии раздражителя приводит к:
Генерацию каких потенциалов нарушит тетраэтиламмоний (ТЭА), блокирующий калиевые каналы?
Как меняется поляризация мембраны под катодом при включении постоянного тока пороговой величины?
происходит частичная деполяризация
+происходит деполяризация и возникает потенциал действия
Как меняется возбудимость при действии сильного длительного постоянного тока:
под катодом — повышается, а под анодом — понижается
+под анодом — повышается, а под катодом — понижается
и под анодом, и под катодом — понижается
Когда и под каким электродом возникает возбуждение при действии порогового постоянного тока:
+при замыкании — под катодом, а при размыкании — под анодом
при замыкании — под анодом, а при размыкании — под катодом
и при замыкании, и размыкании — под катодом
и при замыкании, и размыкании — под анодом
Как меняется возбудимость клетки при действии постоянного тока допороговой величины:
+под катодом — повышается, а под анодом — понижается
под анодом — повышается, а под катодом — понижается
и под катодом, и под анодом — понижается
нигде не изменяется
Лабильность тем выше, чем:
+больше волн возбуждения ткань может воспроизвести в единицу времени
меньше волн возбуждения ткань может воспроизвести в единицу времени
Ткань обладает наибольшей возбудимостью, если ее хронаксия равна:
Ткань обладает наименьшей возбудимостью, если ее хронаксия равна:
Ткань обладает наибольшей лабильностью, если длительность ее потенциалов действия (ПД) равна:
Какая ткань обладает наибольшей возбудимостью, если ее реобаза равна:
Какая ткань обладает наименьшей возбудимостью, если ее реобаза равна:
Ткань обладает наименьшей лабильностью, если длительность ее потенциалов действия (ПД) равна:
Связь между хронаксией и лабильностью:
Нарушит ли процесс генерации потенциала действия (ПД) тетраэтиламмоний (ТЭА), блокирующий калиевые каналы:
Нарушит ли процесс генерации потенциала покоя (ПП) тетраэтиламмоний (ТЭА), блокирующий калиевые каналы:
Генерация каких поте