Сердечная мышца ткань
17. Мышечные ткани. Сердечная и гладкая мышечные ткани
17. Мышечные ткани. Сердечная и гладкая мышечные ткани
Сердечная мышечная ткань
Структурно-функциональной единицей сердечной поперечнополосатой мышечной ткани является кар-диомиоцит. По строению и функциям кардиомиоциты подразделяются на две группы:
1) типичные, или сократительные, кардиомиоциты, образующие своей совокупностью миокард;
2) атипичные кардиомиоциты, составляющие проводящую систему сердца.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку в центре которой локализуется обычно одно ядро.
Атипичные кардиомиоциты образуют проводящую систему сердца, которая включает в себя следующие структурные компоненты:
1) синусо-предсердный узел;
2) предсердно-желудочковый узел;
3) предсердно-желудочковый пучок (пучок Гисса) – ствол, правую и левую ножки;
4) концевые разветвления ножек (волокна Пуркинье). Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
Источники развития кардиомиоцитов – миоэпикар-диальные пластинки, представляющие собой определенные участки висцеральных спланхиотомов.
Гладкая мышечная ткань мезенхимального происхождения
Локализуется в стенках полых органов (желудка, кишечника, дыхательных путей, органов мочеполовой системы) и в стенках кровеносных и лимфатических сосудов. Структурно-функциональной единицей является миоцит: клетка веретенообразной формы длиной 30—100 мкм (в беременной матке – до 500 мкм), диаметром 8 мкм, покрытая базальной пластинкой.
Миозиновые и актиновые филаменты составляют сократительный аппарат миоцита.
Эфферентная иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой.
Сокращение гладкомышечной ткани обычно бывает длительным, что обеспечивает поддержание тонуса полых внутренних органов и сосудов.
Гладкомышечная ткань не образует мышцы в анатомическом понимании этого слова. Однако в полых внутренних органах и в стенке сосудов между пучками миоцитов содержатся прослойки рыхлой волокнистой соединительной ткани, образующие своеобразный эндомизий, а между пластами гладкой мышечной ткани – перимизий.
Регенерация гладкомышечной ткани осуществляется несколькими способами:
1) посредством внутриклеточной регенерации (гипертрофии при усилении функциональной нагрузки);
2) посредством митотического деления миоцитов (пролиферации);
3) посредством дифференцировки из камбиальных элементов (из адвентициальных клеток и миофи-бробластов).
http://med.wikireading.ru/11806
Мышечная ткань: виды, особенности строения и функции
Мышечные ткани — это ткани, отличающиеся по структуре и происхождению, но имеют общую способность к сокращению. Состоят из миоцитов — клеток, которые могут воспринимать нервные импульсы и отвечать на них сокращением.
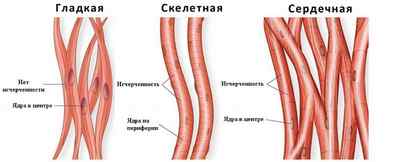
Свойства и виды мышечной ткани
Морфологические признаки:
- Вытянутая форма миоцитов;
- продольно размещены миофибриллы и миофиламенты;
- митохондрии находятся вблизи сократительных элементов;
- присутствуют полисахариды, липиды и миоглобин.
Свойства мышечной ткани:
- Сократимость;
- возбудимость;
- проводимость;
- растяжимость;
- эластичность.
Выделяют следующие виды мышечной ткани в зависимости от морфофункциональных особенностей:
Гистогенетическая классификация делит мышечные ткани на пять видов в зависимости от эмбрионального источника:
- Мезенхимные — десмальный зачаток;
- эпидермальные — кожная эктодерма;
- нейральные — нервная пластинка;
- целомические — спланхнотомы;
- соматические — миотом.
Из 1-3 видов развиваются гладкомышечные ткани, 4, 5 дают поперечнополосатые мышцы.
Строение и функции гладкой мышечной ткани
Cостоит из отдельных мелких веретеновидных клеток. Эти клетки имеют одно ядро и тонкие миофибриллы, которые тянутся от одного конца клетки к другому. Гладкие мышечные клетки объединяются в пучки, состоящие из 10-12 клеток. Это объединение возникает благодаря особенностям иннервации гладкой мускулатуры и облегчает прохождение нервного импульса на всю группу гладких мышечных клеток. Сокращается гладкая мышечная ткань ритмично, медленно и на протяжении длительного времени, способна при этом развивать большую силу без значительных затрат энергии и без утомления.
У низших многоклеточных животных из гладкой мышечной ткани состоят все мышцы, тогда как у позвоночных животных она входит в состав внутренних органов (кроме сердца).
Сокращения этих мышц не зависят от воли человека, т. е. происходят непроизвольно.
Функции гладкой мышечной ткани:
- Поддерживание стабильного давления в полых органах;
- регуляция уровня кровяного давления;
- перистальтика пищеварительного тракта, перемещения по нему содержимого;
- опорожнение мочевого пузыря.
Строение и функции скелетной мышечной ткани
Cостоит из длинных и толстых волокон длиной 10-12 см. Скелетная мускулатура характеризуется произвольным сокращением (в ответ на импульсы, идущие из коры головного мозга). Скорость ее сокращения в 10-25 раз выше, чем в гладкой мышечной ткани.
Мышечное волокно поперечнополосатой ткани покрыто оболочкой — сарколеммой. Под оболочкой находится цитоплазма с большим количеством ядер, расположенных по периферии цитоплазмы, и сократительными нитями — миофибриллами. Состоит миофибрилла из последовательно чередующихся темных и светлых участков (дисков), обладающих разным коэффициентом преломления света. С помощью электронного микроскопа установлено, что миофибрилла состоит из протофибрилл. Тонкие протофибриллы построены из белка — актина, аболее толстые — из миозина.
При сокращении волокон происходит возбуждение сократимых белков, тонкие протофибриллы скользят по толстым. Актин реагирует с миозином, и возникает единая актомиозиновая система.
Функции скелетной мышечной ткани:
- Динамическая — перемещение в пространстве;
- статическая — поддержание определенной позиции частей тела;
- рецепторная — проприорецепторы, воспринимающие раздражение;
- депонирующая — жидкость, минералы, кислород, питательные вещества;
- терморегуляция — расслабление мышц при повышении температуры для расширения сосудов;
- мимика — для передачи эмоций.
Строение и функции сердечной мышечной ткани
Миокард построен из сердечной мышечной и соединительной ткани, с сосудами и нервами. Мышечная ткань относится к поперечнополосатой мускулатуре, исчерченность которой также обусловлена наличием разных типов миофиламентов. Миокард состоит из волокон, которые связаны между собой и формируют сетку. Эти волокна включают одно или двухъядерные клетки, что расположены в виде цепочки. Они получили название сократительных кардиомиоцитов.
Сократительные кардиомиоциты длиной от 50 до 120 микрометров, шириной — до 20 мкм. Ядро здесь располагается в центре цитоплазмы, в отличие от ядер поперечно полосатых волокон. Кардиомиоциты имеют больше саркоплазма и меньше миофибрилл, в сравнении со скелетными мышцами. В клетках сердечной мышцы находится много митохондрий, так как непрерывные сердечные сокращения требуют много энергии.
Вторая разновидность клеток миокарда — это проводящие кардиомиоциты, которые формируют проводящую систему сердца. Проводящие миоциты обеспечивают передачу импульса к сократительным мышечным клеткам.
Функции сердечной мышечной ткани:
- Насосная;
- обеспечивает ток крови в кровеносном русле.
Компоненты сократительной системы
Особенности строения мышечной ткани обусловлены выполняемыми функциями, возможностью принимать и проводить импульсы, способностью к сокращению. Механизм сокращения заключается в согласованной работе ряда элементов: миофибрилл, сократительных белков, митохондрий, миоглобина.
В цитоплазме мышечных клеток имеются особые сократительные нити — миофибриллы, сокращение которых возможно при содружественной работе белков — актина и миозина, а также при участии ионов Са. Митохондрии снабжают все процессы энергией. Также энергетические запасы образуют гликоген и липиды. Миоглобин необходим для связывания O2 и формирование его запаса на период сокращения мышцы, так как во время сокращения идет сдавление кровеносных сосудов и снабжение мышц O2 резко снижается.
Таблица. Соответствие между характеристикой мышечной ткани и ее видом
http://animals-world.ru/myshechnaya-tkan/
Сердечная мышечная ткань
Структурными единицами сердечной мышечной ткани являются клетки – кардиомиоциты, покрытые базальной мембраной.
Различают 5 видов кардиомиоцитов: сократительные (рабочие), или типичные, и атипичные: синусные (пейсмекерные), переходные, проводящие и секреторные.
Рабочие кардиомиоциты имеют форму удлинённого цилиндра длиной около 100-150 мкм и диаметром до 20 мкм. Они содержат одно, реже два ядра, располагающихся по центру клетки, а вокруг ядер группами локализуются миофибриллы (поля Конгейма). Строение миофибрилл такое же, как и в скелетной мышечной ткани, но в них отсутствуют триады. Кардиомиоциты соединяются торец в торец, образуя функциональные мышечные волокна. В области соединений кардиомиоцитов на светооптическом уровне чётко выявляются вставочные диски.
Во Вставочных дисках различают продольные и поперечные участки:
В Поперечных участках имеется много межклеточных контактов — Десмосом, они обеспечивают прочность соединения кардиомиоцитов; в Продольных Участках присутствует много межклеточных контактов типа Нексусов, которые образуют узкие каналы между соседними клетками, через эти каналы способна проходить вода и ионы, что создает условия для свободного прохождения электрического тока с одного кардиомиоцита на другой; таким образом, наличие нексусов обеспечивает электрическое сопряжение кардиомиоцитов, необходимое для быстрого распространения возбуждения по всей массе миокарда и для его синхронного сокращения
Пейсмекерные кардиомиоциты (Р-клетки) располагаются в области синусов. Они способны ритмически сокращаться и передавать управляющие сигналы через переходные и проводящие кардиомиоциты на рабочие, которые сокращаются с заданным ритмом.
Переходные и Проводящие кардиомиоциты Передают возбуждение сердечного ритма от Р-клеток к сократительным кардиомиоцитам.
Секреторные кардиомиоциты Производят предсердный натрийуретичний фактор, который регулирует мочеобразование, является антогонистом ренина (усиливает диурез и снижает артериальное давление).
Общим для морфологии скелетной и сердечной мышечной тканей является наличие исчерченности, выявляемой на светооптическом уровне, и, так называемых, Т-трубочек, обнаруживаемых при ультрамикроскопическом исследовании.
Т-трубочки — это трубкообразные впячивания цитомембраны, идущие внутрь мышечного волокна и кардиомиоцита, то есть располагаются поперечно относительно их длины. Примерно на уровне Z-линий, они подходят близко к эндоплазматическому ретикулуму.
Гладкая мышечная ткань
В гладкой мышечной ткани мезенхимного генеза структурной единицей является миоцит, имеющий веретеновидную форму, ядро в нём удлинённое, локализуется по центру клетки. Длина миоцитов колеблется в пределах 20-500 мкм, а ширина в области брюшка – всего лишь 5-8 мкм. Сократительный аппарат представлен филаментами актина, образующими трёхмерную сеть, рядом с которой располагаются мономеры миозина.
В гладкой мышечной ткани тропонин-тропомиозинового комплекса нет, головка миозина имеет легкие цепи, которые должны сначала фосфорилироваться, для того чтобы она смогла расщеплять и присоединять АТФ и взаимодействовать с актином.
Структурной единицей гладких мышц эктодермального происхождения является миоэпителиоцит экзокринных желез, а нейрального – мионейральные клетки m. m. sphincter et dilatator pupille.
http://veterinarua.ru/tkani/86-serdechnaya-myshechnaya-tkan.html
Сердечная мышечная ткань;
Сердечная мышечная ткань формирует среднюю оболочку (миокард) предсердий и желудочков сердца и представлена двумя разновидностями рабочей и проводящей.
Рабочая мышечная ткань состоит из клеток кардиомиоцитов, важнейшей особенностью которых является наличие совершенных контактных зон. Соединяясь друг с другом, торцевыми концами они формируют структуру, сходную с мышечным волокном. На боковых поверхностях кардиомиоциты имеют ответвления. Соединяясь концами с ответвлениями соседних кардиомиоцитов они образуют анастомозы. Границами между торцами соседних кардиомиоцитов являются вставочные диски с прямыми или ступенчатыми контурами. В световом микроскопе они имеют вид поперечных темных полосок. С помощью вставочных дисков и анастомозов сформирована единая структурно-функциональная сократительная система.
При электронной микроскопии выявлено, что в области вставочных дисков одна клетка вдается в другую пальцевидными выступами, на боковых поверхностях которых имеются десмосомы, что обеспечивает высокую прочность сцепления. На концах пальцевидных выступов обнаружены щелевидные контакты, через которые нервные импульсы быстро распространяются от клетки к клетке без участия медиатора синхронизируя сокращение кардиомиоцитов.
Сердечные миоциты – это одноядерные, иногда двухядерные клетки. Ядра расположены в центре в отличие от скелетных мышечных волокон. В околоядерной зоне расположены компоненты аппарата Гольджи, митохондрии, лизосомы, гранулы гликогена.
Сократительный аппарат миоцитов, так же как и в скелетной мышечной ткани, состоит из миофибрилл, которые занимают периферическую часть клетки. Их диаметр от 1 до 3-х мкм.
Миофибриллы сходны с миофибриллами скелетной мышечной ткани. Они также построены из анизотропных и изотропных дисков, что также обуславливает поперечную исчерченность.
Плазмолемма кардиомиоцитов на уровне Z-полосок инвагинирует в глубь цитоплазмы, образуя поперечные трубочки, отличающиеся от скелетной мышечной ткани большим диаметром и наличием базальной мембраны, которая покрывает их снаружи, как и сарколемму. Волны деполяризации, идущие с плазмолеммы внутрь сердечных миоцитов, вызывают скольжение актиновых миофиламентов (протофибрилл) по отношению миозиновым, обуславливая сокращение, как и в скелетной мышечной ткани.
Т-трубочки в сердечных рабочих кардиомиоцитах образуют диады, то есть связаны с цистернами саркоплазматической сети только с одной стороны. Рабочие кардиомиоциты имеют длину 50-120 мкм, ширину 15-20 мкм. Количество миофибрилл в них меньше, чем в мышечных волокнах.
Сердечная мышечная ткань содержит много миоглобина, поэтому темно-красного цвета. В миоцитах много митохондрий и гликогена, т.е.: энергию сердечная мышечная ткань получает и при распаде АТФ, и в результате гликолиза. Таким образом, сердечная мышца работает непрерывно всю жизнь, из-за мощной энергетической оснащенности.
Интенсивность и частота сокращений сердечной мышцы регулируются нервными импульсами.
В эмбриогенезе рабочая мышечная ткань развивается из особых участков висцерального листка несегментированной мезодермы (спланхнотома). В сформировавшейся рабочей мышечной ткани сердца отсутствуют камбиальные клетки (миосателлиты), поэтому при повреждении миокарда в травмированной зоне кардиомиоциты погибают и на месте повреждения развивается волокнистая соединительная ткань.
Проводящая мышечная ткань сердца находится в составе комплекса образований синусно-предсердного узла, расположенного в устье краниальной полой вены, предсердно-желудочкового узла, лежащего в межпредсердной перегородке, предсердно-желудочкового ствола (пучка Гиса) и его разветвлений, находящихся под эндокардом межжелудочковой перегородки и в соединительно-тканных прослойках миокарда.
Все компоненты этой системы образованы атипичными клетками, специализированными либо на выработке импульса, распространяющемуся по всему сердцу и вызывающего сокращение его отделов в необходимой последовательности (ритме), либо в проведении импульса к рабочим кардиомиоцитам.
Для атипичных миоцитов характерен значительный объем цитоплазмы, в которой немногочисленные миофибриллы занимают периферическую часть и не имеют параллельной ориентации, вследствие чего этим клеткам не свойственна поперечная исчерченность. Ядра расположены в центре клеток. Цитоплазма богата гликогеном, но в ней мало митохондрий, что свидетельствует об интенсивном гликолизе и низком уровне аэробного окисления. Поэтому клетки проводящей системы более устойчивы к кислородному голоданию, чем сократительные кардиомиоциты.
В составе синусно-предсердного узла атипичные кардиомиоциты более мелкие, округлой формы. В них формируются нервные импульсы и они относятся к главным водителям ритма. Миоциты предсердно-желудочкового узла несколько крупнее, а волокна пучка Гиса (волокна Пуркинье) состоят из крупных округлых и овальных миоцитов с эксцентрично расположенным ядром. Диаметр их в 2-3 раза больше, чем рабочих кардиомиоцитов. Электронно-микроскопически выявлено, что в атипичных миацитах слаборазвита саркоплазматическая сеть, отсутствует система Т-трубочек. Клетки соединяются не только концами, но и боковыми поверхностями. Вставочные диски устроены более просто и не содержат пальцевидных соединений, десмосом и нексусов.
Гладкая мышечная ткань образует мышечную оболочку трубкообразных органов пищеварения, дыхания, выделения, размножения, находится в стенках кровеносных сосудов, протоков желез, в селезенке, коже и других органах.
Специализированные сократительные гладкомышечные ткани входят в состав потовых, слюнных, молочных желез. Сократительные клетки этих желез в своей цитоплазме содержат миофиламенты, построенные из сократительных белков и развиваются из эпителиальных клеток. Другие разновидности специализированных сократительных тканей имеют нейроглиальное происхождение, суживают зрачок и располагаются в радужной оболочке глаза.
Гладкая мышечная ткань относится к ткани с непроизвольным сокращением, её функцию контролирует вегетативная нервная система. Сокращения гладких мышц могут быть медленными, но достигать большой силы сжатия.
Основной структурной единицей гладкой мышечной ткани являются клетки-миоциты. Они удлиненной веретеновидной формы с заостренными концами. Их длина от 20 до 200 мкм (в беременной матке до 500 мкм), а толщина 8-10 мкм. Ядро палочковидной формы находится в середине клетки. В цитоплазме, около полюсов ядра расположены органеллы: митохондрии, комплекс Гольджи, центросома, рибосомы, эндоплазматическая сеть и включения гликогена (энергетический резерв клетки). В преферической части цитоплазмы расположены миофиламенты. Нити актина и миозина не образуют миофибрилл или постоянных акто-миозиновых комплексов и расположены по-разному. Актиновые нити чаще имеют косое присоединение к плазмолемме с помощью особых плотных телец (прикрепительных дисков). Отдельные пучки актиновых нитей прикрепляются к плотным тельцам, расположенным в цитоплазме. Положение этих телец или дисков с обратной стороны закрепляется промежуточными филаментами.
Миозиновые нити в периоды расслабления миоцитов лежат в цитоплазме продольно или под углом к длинной оси клетки. В процессе сокращения актиновые и миозиновые нити смещаются навстречу друг другу и формируют акто-миозиновые комплексы. В результате клетка сокращается и приобретает неправильную форму. В фазе расслабления комплексы вновь распадаются. Поскольку актиновые и миозиновые нити лежат неупорядоченно, поперечная исчерченность в гладких миоцитах отсутствует.
В процессе сокращения, как было сказано, важную роль играют ионы Са ++ . Депо для них является гладкая эндоплазматическая сеть миоцита. Кроме того, ионы Са ++ поступают извне через кальциевые каналы в цитолемме. В определенных участках плазмолеммы лежат специальные белки, воспринимающие и пропускающие внутрь ионы Са ++ . Ионы Са ++ в комплексе с белком кольмодулином и ферментом киназой запускают процесс сокращения. Головки молекул миозина начинают двигаться и скользить вдоль нитей актина и осуществляется процесс сокращения.
С помощью электронного микроскопа было выявлено, что на концах гладких миоцитов имеются пальцевидные выпячивания, десмосомы и щелевидные контакты-нексусы. Плазмолемма миоцитов впячиваясь в цитоплазму, образует пузырьки (кавеолы), примыкающие к саркоплазматической сети. Предполагают, что эти пузырьки участвуют в проведении нервных импульсов, вызывающих выход ионов Са ++ и процесс сокращения.
Функциональной единицей гладкой мышечной ткани является пучок из 10-15 миоцитов, связанных с одним нервным волокном. Благодаря тесной связи клеток с помощью десмосом и щелевых контактов все клетки пучка быстро реагируют на нервное раздражение, несмотря на то, что нервное окончание входит только в одну клетку.
Коллагеновые волокна, соединительно-тканных капсул (эндомизий) оплетают миоциты, вплетаются в базальную пластинку (мембрану) в наружный слой над сарколеммой и тем самым удерживают клетки от чрезмерного сжатия и растяжения.
Пучки отделены друг от друга прослойками соединительной ткани (перемизий), в которой проходят сосуды и нервы.
Гладкая мышечная ткань, иннервируется вегетативной нервной системой. Ее деятельность регулируется корой полушарий, но без участия сознания. Сокращения осуществляются непроизвольно и происходят медленно и ритмично (период сокращения от 3 до 5 минут).
Такой характер сокращения называют тоническим.
В стенках полых органов и сосудов пучки гладких миоцитов объединяются в пласты (продольные и циркулярные).
Гладкая мышечная ткань обладает большой силой, передвигая в кишечнике большие массы пищи, и обладает слабой утомляемостью. В стенке кишечника сокращение происходит 12 раз в минуту.
Отдельные пучки гладких миоцитов находятся в коже животных в виде мышц, поднимающих волос.
Происходит гладкая мышечная ткань внутренних органов и сосудов из мезенхимы, клетки которой дифференцируются в миобласты, а миобласты в миоциты, сохраняющие способность к делению на протяжении всего онтогенеза. Кроме того, гладкие миоциты могут образовываться из недеффиренцированных клеток соединительной ткани (адвентициальных), находящихся около кровеносных сосудов.
http://studopedia.su/10_117627_serdechnaya-mishechnaya-tkan.html
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Эта ткань образует один из слоев стенки сердца — миокард. Она делится на собственно сердечную мышечную ткань и проводящую систему.
Собственно сердечная мышечная ткань по своим физиологическим свойствам занимает промежуточное положение между гладкими мышцами внутренних органов и поперечнополосатыми (скелетными).
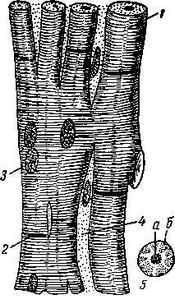
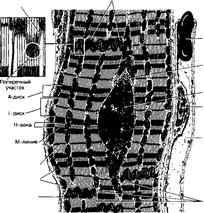
Рис. 66. Схема строения
/ — мышечное волокно; 2 — вставочные диски; 3 — ядро; 4 -— прослойка рыхлой соединительной ткани; 5 — поперечный разрез мышечного волокна; а — ядро; б — пучки миофибрилл, расположен* ные по радиусам.
быстрее гладких, но медленнее поперечнополосатых мышц, работает ритмично и мало утомляется. В связи с этим в ее строении имеется ряд своеобразных черт (рис. 66). Состоит эта ткань из отдельных мышечных клеток (миоцитов), почти прямоугольной формы, расположенных столбиком друг за другом. В целом получается структура, напоминающая поперечнополосатое волокно, разделенное на отрезки поперечными перегородками — вставочные диски, являющиеся участками плазмалеммы двух соседних клеток, соприкасающихся друг с другом. Рядом лежащие волокна соединены анастомозами, что позволяет им сокращаться одновременно. Группы мышечных волокон окружены соединительнотканными прослойками, подобными эндоми-зию. В центре каждой клетки 1—2 ядра овальной формы. Миофибриллы располагаются по периферии клетки и имеют поперечную исчер-ченность. Между миофибриллами в саркоплазме большое количество митохондрий (саркосом), чрезвычайно богатых кристами, что говорит о высокой их энергетической активности. Снаружи клетка покрыта, кроме плазмалеммы, еще и базальной мембраной. Богатство цитоплазмой и хорошо развитый трофический аппарат обеспечивают сердечной мышце непрерывность деятельности.
Проводящая система сердца состоит из бедных миофибриллами тяжей мышечной ткани»способных согласовывать работу разобщенных мышц желудочков и предсердий.
http://xn--80ahc0abogjs.com/veterinariya_727/serdechnaya-myishechnaya-tkan.html
Сердечная мышечная ткань: источник развития, структурная и функциональная характеристика ткани, особенности иннервация и сократительной активности, виды кардиомиоцитов, регенерация.
сердечная мышечная ткань как разновидность поперечнополосатой мышечной ткани имеет общую функцию и ряд структурных признаков, сходных со скелетной поперечнополосатой мышечной тканью. Организация миофибрилл и механизм сокращения одинаковы. Вместе с тем, сердечная мышечная ткань обладает рядом отличий, которые суммированы в таблице 1.
Таблица 1. Сравнительная характеристика скелетной и сердечной поперечнополосатой мышечной ткани.
Скелетная мышечная ткань
Сердечная мышечная ткань
Висцеральный листок спланхнотома
Мышечное волокно (симпласт и миосателлитоциты)
На периферии мио-симпласта – множество
В центре кардиомиоцита – одно, иногда — два
Локализация сократительного аппарата
В центре миосимпласта
На периферии кардио-миоцита
Наличие белых, красных и промежуточных мышечных волокон. Наличие камбия – миосателлитоцитов
Наличие сократитель-ных, мышечно-секре-торных и проводящих кардиомицитов.
Наличие вставочных дисков и анастомозов
Соматическая нервная система
Вегетативная нервная система
Участие в терморегуляции и углеводном обмене
Синтез гормонов (атриопептидов)
Физиологическая и репаративная (за счет миосателлитоцитов)
Физиологическая. Погибшие кардиомиоциты не восстанавливаются
Кардиомиоцит
Структурно-функциональной единицей является клетка —кардиомиоцит.
Классификация кардиомиоцитов
По строению и функциям кардиомиоциты подразделяются на две основные группы:
типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит
представляет собой почти прямоугольную клетку 50—120 мкм в длину, шириной 15—20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой.
В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии.
В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах.
Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть — функциональный синтиций.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (диском между кардиомиоцитами нет.
Вставочные диски
— это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты.
В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты.
Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Атипичные кардиомиоциты
образуют проводящую систему сердца, состоящую из:
предсердно-желудочковый пучок (пучок Гиса)ствол, правую и левую ножки;
концевые разветвления ножек — волокна Пункинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичным рядом особенностей:
они крупнее (длина 100 мкм, толщина 50 мкм);
в цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
плазмолемма не образует Т-канальцев;
во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
Р-клетки (пейсмекеры) водители ритма (I типа);
переходные клетки (II типа);
клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов — миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнееиз целомического эпителия этих участков.
Иннервация сердечной мышечной ткани
Биопотенциалы сократительные кардиомиоциты получают из двух источников:
из проводящей системы сердца (прежде всего из синусо-предсердного узла);
из вегетативной нервной системы (из ее симпатической и парасимпатической части).
Регенерация сердечной мышечной ткани
Кардиомиоциты регенерируют только по внутриклеточному типу. Пролиферации кардиомиоцитов не наблюдается. Камбиальные элементы в сердечной мышечной ткани отсутствуют. При поражении значительных участков миокарда (в частности, при инфаркте миокарда) восстановление дефекта происходит за счет разрастания соединительной ткани и образования рубцов (пластическая регенерация). Естественно, что сократительная функция в этих участках отсутствует. Поражение проводящей системы сопровождается нарушением ритма сердечных сокращений.
Сердечная мышечная ткань
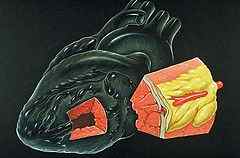
Миокард (лат. myocardium от др.-греч. ??? «мышца» + ?????? «сердце») — название мышечного среднего слоя сердца, составляющего основную часть его массы.
Содержание
Миокард образован сердечной исчерченной поперечнополосатой мышечной тканью, представляющей собой плотное соединение мышечных клеток — кардиомиоцитов, образующих основную часть миокарда. Отличается от других типов мышечной ткани (скелетная мускулатура, гладкая мускулатура) особым гистологическим строением, облегчающим распространение потенциала действия между кардиомиоцитами. Элементарной сократительной единицей кардиомиоцита является саркомер — участок миофибриллы между двумя так называемыми линиями Z. Длина саркомера равна 1,6—2,2 мкм в зависимости от степени сокращения. В саркомере чередуются светлые и тёмные полосы, отчего миофибрилла при световой микроскопии выглядит поперечно исчерченной. В центре находится тёмная полоса постоянной длины (1,5 мкм) — диск A, его ограничивают два более светлых диска I переменной длины. Саркомер миокарда, как и скелетной мышцы, состоит из переплетённых нитей (миофиламентов) двух типов. Толстые нити есть только в диске A. Они состоят из белка миозина, имеют сигарообразную форму, диаметр 10 нм и длину 1,5—1,6 мкм. Тонкие нити включают прежде всего актин и идут от линии Z через диск I в диск A. Их диаметр составляет 5 нм, длина — 1 мкм. Толстые и тонкие нити накладываются друг на друга только в диске A; диск I содержит лишь тонкие нити. При электронной микроскопии между толстыми и тонкими нитями видны поперечные мостики.
Особенности [ | ]
Главная особенность миокарда — создание ритмических движений сердца. Функциональной особенностью миокарда являются ритмичные автоматические сокращения, чередующиеся с расслаблениями, совершаются непрерывно в течение всей жизни организма. Последовательное сокращение и расслабление различных отделов сердца связано с его строением и наличием проводящей системы сердца, по которой распространяется импульс. Миокард предсердий и желудочков разобщён фиброзной перегородкой, что позволяет им сокращаться независимо друг от друга, так как возбуждение не может распространяться по фиброзной ткани. Возбуждение от предсердий к желудочкам проводится только через атриовентрикулярный пучок, отходящий от атриовентрикулярного узла [1] .
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
РАЗВИТИЕ. Источником развития сердечной мьшючной ткани является миоэпикардиальная пластинка— часть висцерального сплаихпотома в шейном отделе зародыша. Ее клетки превращаются в миобласты, которые активно делятся митозом и дифференцируются. В цитоплазме миобластов синтезируются миофиламенты, формирующие миофибриллы. Вначале миофибриллы не имеют исчерченности и определенной ориентации в цитоплазме. В процессе дальнейшей дифференцировки принимают продольную ориентацию и тонкими миофиламентами прикрепляются к формирующимся уплотнениям сарколеммы (Z-вещество).
В результате все возрастающей упорядоченности миофиламентов миофибриллы приобретают поперечную исчерчениость. Образуются кардиоми- оциты. В их цитоплазме нарастает содержание органелл: митохондрий, гра нулярной ЭПС, свободных рибосом. В процессе дифференцировки кардио миоциты не сразу теряют способность к делению и продолжают размножаться. В некоторых клетках может отсутствовать цитотомия, что ведет к появлению двуядерных кардиомиоцитов. Развивающиеся кардиомиоциты имеют строго определенную пространственную ориентацию, выстраиваясь в виде цепочек и образуя друг с другом межклеточные контакты — вставочные диски. В результате дивергентной дифференцировки кардиомиоциты превращаются в клетки трех типов: 1) рабочие, или типичные, сократительные; 2) проводящие, или атипичные; 3) секреторные (эндокринные). В результате терминальной дифференцировки кардиомиоциты к моменту рождения или в первые месяцы постнаталыюго онтогенеза теряют способность к делению. В зрелой сердечной мышечной ткани камбиальные клетки отсутствуют.
СТРОЕНИЕ. Сердечная мышечная ткань образована клетками карди-омиоцитами. Кардиомиоциты являются единственным тканевым элементом сердечной мышечной ткани. Они соединяются друг с другом при помощи вставочных дисков и образуют функциональные мышечные волокна, или функциональный симпласт, не являющийся симпластом в морфологическом понятии. Функциональные волокна разветвляются и анастомози-руют боковыми поверхностями, в результате чего образуется сложная трехмерная сеть (рис. 12.15).
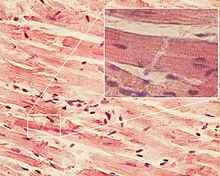
Кардиомиоциты имеют вытянутую прямоугольную слабоотростчатую форму. Они состоят из ядра и цитоплазмы. Многие клетки (более половины у взрослого индивидуума) являются двуядерными и полиплоидными. Степень полиплоидизации различна и отражает адаптивные возможности миокарда. Ядра крупные, светлые, находятся в центре кардиомиоцитов.
Цитоплазма (саркоплазма) кардиомиоцитов обладает выраженной ок-сифилией. В ней содержится большое количество органелл и включений. Периферическую часть саркоплазмы занимают расположенные продольно исчерченные миофибриллы, построенные так же, как в скелетной мышечной ткани (рис. 12.16). В отличие от миофибрилл скелетной мышечной ткани, лежащих строго изолированно, в кардиомиоцитах миофибриллы нередко сливаются друг с другом с образованием единой структуры и содержат сократимые белки, химически отличающиеся от сократимых белков миофибрилл скелетных мышц.
СИР и Т-трубочки развиты слабее, чем в скелетной мышечной ткани, что связано с автоматией сердечной мышцы и меньшим влиянием нервной системы. В отличие от скелетной мышечной ткани СПР и Т-трубочки образуют не триады, а диады (к Т-трубочке прилежит одна цистерна СПР). Типичные терминальные цистерны отсутствуют. СПР менее интенсивно аккумулирует кальций. Снаружи кардиоциты покрыты сарколеммой, состоящей из плаз-молеммы кардиомпоцита и базаль-ной мембраны снаружи. Вазальная мембрана тесно связана с межклеточным веществом, в нес вплетаются коллагеновые и эластические волокна. Базальная мембрана отсутствует в местах вставочных дисков. Со вставочными дисками связаны компоненты цитоскелета. Через интегрины цитолеммы они также связаны с межклеточным веществом. Вставочные диски — это место контактов двух кардиомиоцитов, комплексы межклеточных контактов. Они обеспечивают как механическую, так и химическую, функциональную коммуникацию кардиомиоцитов. В световом микроскопе имеют вид темных поперечных полосок (рис. 12.14 б). В электронном микроскопе вставочные диски имеют зигзагообразный, ступеньчатый вид или вид зубчатой линии. В них можно выделить горизонтальные и вертикальные участки и три зоны (рис. 12.1,12.15 6).
1. Зоны десмосом и полосок слипания. Находятся на вертикальных (поперечных) участках дисков. Обеспечивают механическое соединение кардиомиоцитов.
2. Зоны нексусов (щелевых контактов) — места передачи возбуждения с одной клетки на другую, обеспечивают химическую коммуникацию кардиомиоцитов. Обнаруживаются на продольных участках вставочных дисков.3. Зоны прикрепления миофибрилл.Находятся на поперечных участках вставочных дисков. Служат местами прикрепления актиновых фила-ментов к сарколемме кардиоми-оцита. Это прикрепление происходит к Z-полоскам, обнаруживаемым на внутренней поверхности сарколеммы и аналогичным Z-линиям. В области вставочных дисков обнаруживаются в большом количестве кадгерины(адгезивные молекулы, осуществляющие каль-цийзависимую адгезию кардио-миоцитов друг с другом).
Типы кардиомиоцитов.Кардиомиоциты имеют разные свойства в разных участках сердца. Так, в предсердиях они могут делиться митозом, а в желудочках никогда не делятся. Различают три тина кардиомиоцитов, существенно отличающихся друг от друга гак строением, так и функциями: рабочие, секреторные, проводящие.
1. Рабочие кардиомиоцитыимеют структуру, описанную выше.
2. Среди предсердных миоцитов есть секреторные кардиомиоциты,которые вырабатывают натрийуретический фактор (НУФ),усиливающий секрецию натрия почками. Кроме этого, НУФ расслабляет гладкие ми-оциты стенки артерий и подавляет секрецию гормонов, вызывающих гипер-тензию (альдостеронаи вазопрессина).Все это ведет к увеличению диуреза и просвета артерий, снижению объема циркулирующей жидкости и в результате — к снижению артериального давления. Секреторные кардиомиоциты локализуются в основном в правом предсердии. Следует отметить, что в эмбриогенезе способностью к синтезу обладают все кардиомиоциты, но в процессе дифференцировки кардиомиоциты желудочков обратимо те-ряют эту способность, которая может восстанавливаться здесь при перенапряжении сердечной мышцы.

3. Значительно отличаются от рабочих кардиомиоцитов проводящие (атипичные) кардиомиоциты.Образуют проводящую систему сердца (см. \»сердечно-сосудистую систему\»). Они в два раза больше рабочих кардиомиоцитов. В этих клетках содержится мало миофибрилл, увеличен объем саркоплазмы, в которой выявляется значительное количество гликогена. Благодаря содержанию последнего цитоплазма атипичных кардиомиоцитов плохо воспринимает окраску. В клетках содержится много лизосом и отсутствуют Т-трубочки. Функцией атипичных кардиомиоцитов является генерация электрических импульсов и передача их на рабочие клетки. Несмотря на автоматизм, работа сердечной мышечной ткани строго регулируется вегетативной нервной системой. Симпатическая нервная система учащает и усиливает, парасимпатическая — урежает и ослабляет сердечные сокращения.
РЕГЕНЕРАЦИЯ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ. Физиологическая регенерация.Реализуется на внутриклеточном уровне и протекает с высокой интенсивностью и скоростью, поскольку сердечная мышца несет огромную нагрузку. Еще более она возрастает при тяжелой физической работе и в патологических условиях (гипертоническая болезнь и др.). При этом происходит постоянное изнашивание компонентов цитоплазмы кардиомиоцитов и замещение их вновь образованными. При повышенной нагрузке на сердце происходит гипертрофия(увеличение размеров) и гиперплазия(увеличение количества) органелл, в том числе и миофибрилл с нарастанием в последних количества саркомеров. В молодом возрасте отмечаются также полиплоидизация кардиомиоцитов и появление двуядерных клеток. Рабочая гипертрофия миокарда характеризуется адекватным адаптивным разрастанием его сосудистого русла. При патологиии (например, пороки сердца, также вызывающие гипертрофию кардиомиоцитов) этого не происходит, и через некоторое время из-за нарушения питания происходит гибель части кардиомиоцитов с замещением их рубцовой тканью (кардиосклероз).
Репаративная регенерация.Происходит при ранениях сердечной мышцы, инфарктах миокарда и при других ситуациях. Поскольку в сердечной мышечной ткани пет камбиальных клеток, то при повреждении миокарда желудочков регенераторные и адаптивные процессы идут на внутриклеточном уровне в соседних кардиомиоцитах: они увеличиваются в размерах и берут на себя функцию погибших клеток. На месте погибших кардиомиоцитов образуется соединительнотканный рубец. В последнее время установлено, что некроз кардиомиоцитов при инфаркте миокарда захватывает только кардиомиоциты сравнительно небольшого участка зоны инфаркта и близлежащей зоны. Более значительное количество кардиомиоцитов, окружающих зону инфаркта, погибает путем апрптоза, и этот процесс является ведущим в гибели клеток сердечной мышцы. Поэтому лечение инфаркта миокарда в первую очередь должно быть направлено на подавление апоптоза кардиомиоцитов в первые сутки после наступления инфаркта.
При повреждении миокарда предсердий в небольшом объеме может осуществляться регенерация на клеточном уровне.
Стимуляция репаративной регенерации сердечной мышечной ткани. 1)Предотвращение апоптоза кардиомиоцитов назначением препаратов, улучшающих микроциркуляцию миокарда, снижающих свертывание крови, ее вязкость и улучшающих реологические свойства крови. Успешная борьба с постинфарктным апоптозом кардиомиоцитов является важным условием дальнейшей успешной регенерации миокарда; 2) Назначение анаболических препаратов (витаминного комплекса, препаратов РНК и ДНК, АТФ и др.); 3) Раннее применение дозированных физических нагрузок, комплекса упражнений лечебной физкультуры.
В последние годы в экспериментальных условиях для стимуляции регенерации сердечной мышечной ткани стали применять трансплантацию миосателлитоцитов скелетной мышечной ткани. Установлено, что введенные в миокард миосателлитоциты формируют скелетные мышечные волокна, устанавливающие тесную не только структурную, но и функциональную связь с кардиомиоцитами. Поскольку замещение дефекта миокарда не инертной соединительной, а проявляющей сократительную активность скелетной мышечной тканью более выигрышно в функциональном и даже в механическом отношении, то дальнейшая разработка этого метода может оказаться перспективной при лечении инфарктов миокарда у людей.
http://megaobuchalka.ru/4/39118.html
Под ред. Э.Г. Улумбекова, Ю.А. Челышева, Гистология (введение в патологию), 1997
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Поперечнополосатая мышечная ткань сердечного типа входит в состав мышечной стенки сердца (миокард). Основной гистологический элемент — кардиомиоцит. Кардиомиоциты присутствуют также в проксимальной части аорты и верхней полой вены.
А. Кардиомиогенез. Миобласты происходят из клеток спланхнической мезодермы, окружающей эндокардиальную трубку (глава 10 Б I). После ряда митотических делений G,-mho6- ласты начинают синтез сократительных и вспомогательных белков и через стадию G0- миобластов дифференцируются в кардиомиоциты, приобретая вытянутую форму; в саркоплазме начинается сборка миофибрилл. В отличие от поперечнополосатой мышечной ткани скелетного типа, в кардиомиогенезе не происходит обособления камбиального резерва, а все кардиомиоциты необратимо находятся в фазе G0 клеточного цикла. Специфический фактор транскрипции (ген CATFl/SMBP2, 600502, Ilql3.2-ql3.4) экспрессируется только в развивающемся и сформировавшемся миокарде.
Б. Кардиомиоциты расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры бассейна венечных сосудов и терминальные ветвления двигательных аксонов нервных клеток вегетативного отдела нервной системы. Каждый миоцит имеет сарколемму (базальная мембрана + плазмолемма). Различают рабочие, атипичные и секреторные кардиомиоциты.
а. Сократительный аппарат. Организация миофибрилл и саркомеров в кардиомио- цитах такая же, как и в скелетном мышечном волокне (см. I Б I, 2). Одинаков и механизм взаимодействия тонких и толстых нитей при сокращении (см. I Г 5, 6, 7).
б. Саркоплазматическая сеть. Выброс Ca2+ из саркоплазматического ретикулума регулируется через рецепторы рианодина (см. также главу 2 III А 3 б (3) (а)). Изменения мембранного потенциала открывают потенциалзависимые Са2+-каналы, в кар- диомиоцитах незначительно повышается концентрация Ca2+. Этот Ca2+ активирует рецепторы рианодина, и Ca2* выходит в цитозоль (кальций-индуцированная мобилизация Ca2+).
в. Т-трубочки в кардиомиоцитах, в отличие от скелетных мышечных волокон, проходят на уровне Z-линий. В связи с этим Т-трубочка контактирует только с одной терминальной цистерной. В результате вместо триад скелетного мышечного волокна формируются диады.
г. Митохондрии расположены параллельными рядами между миофибриллами. Их более плотные скопления наблюдают на уровне I-дисков и ядер.
Комплекс Г ольджи
Ядро
Эндотелиальная
клетка
Рис. 7-11. Рабочий кардиомиоцит — удлинённой формы клетка. Ядро расположено центрально, вблизи ядра находятся комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Вставочные диски (на врезке) служат для скрепления кардиомиоцитов и синхронизации их сокращения [из Hees H, Sinowatz F (1992) и Kopf-MaierP, Merker H-J <1989))
д. Вставочные диски. На концах контактирующих кардиомиоцитов имеются интердигитации (пальцевидные выпячивания и углубления). Вырост одной клетки плотно входит в углубление другой. На конце такого выступа (поперечный участок вставочного диска) сконцентрированы контакты двух типов: десмосомы и промежуточные. На боковой поверхности выступа (продольный участок вставочного диска) имеется множество щелевых контактов (nexus, нексус).
е. Предсердные и желудочковые миоциты — разные популяции рабочих кардиомиоцитов. В предсердных кардиомиоцитах слабее развита система Т-трубочек, но в зоне вставочных дисков значительно больше щелевых контактов. Желудочковые кардиомиоциты крупнее, они имеют хорошо развитую систему Т-трубочек. В состав сократительного аппарата миоцитов предсердий и желудочков входят разные изоформы миозина, актина и других контрактильных белков.
а. Водители ритма (пейсмейкерные клетки, пейсмейкеры; рис. 7-12) — совокупность специализированных кардиомиоцитов в виде тонких волокон, окружённых рыхлой соединительной тканью. По сравнению с рабочими кардиомиоцитами они имеют меньшие размеры. В саркоплазме содержится сравнительно мало гликогена и небольшое количество миофибрилл, лежащих в основном по периферии клеток. Эти клетки имеют богатую васкуляризацию и двигательную вегетативную иннервацию. Так, в синусно- предсердном узле доля соединительнотканных элементов (включая кровеносные капилляры) в 1,5-3 раза, а нервных элементов (нейроны и двигательные нервные окончания) в 2,5-5 раз выше, чем в рабочем миокарде правого предсердия. Главное свойство водителей ритма — спонтанная деполяризация плазматической мембраны. При достижении критического значения возникает потенциал действия, распространяющийся по волокнам проводящей системы сердца и достигающий рабочих кардиомиоцитов. Главный водитель ритма — клетки синусно-предсердного узла — генерирует ритм 60-90 импульсов в минуту. Нормально активность других водителей ритма подавлена.
б. Проводящие кардиомиоциты — специализированные клетки, выполняющие функцию проведения возбуждения от водителей ритма. Эти клетки образуют длинные волокна.
В. Иннервация. На деятельность сердца — сложной авторегуляторной и регулируемой системы — оказывает влияние множество факторов, в т.ч. двигательная вегетативная
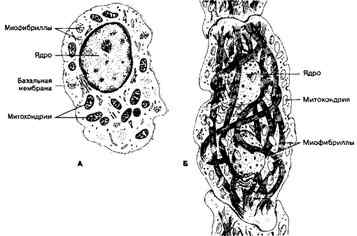
Рис. 7-12. Атипичные кардиомиоциты. А — водитель ритма синусно-предсердного узла;
Б — проводящий кардиомиоцит пучка Гйса [из Hees Н, Sinowatz F, 1992]
иннервация — парасимпатическая и симпатическая. Парасимпатическая иннервация осуществляется терминальными варикозными окончаниями аксонов блуждающего нерва, а симпатическая — окончаниями аксонов адренергических нейронов шейного верхнего, шейного среднего и звёздчатого (шейно-грудного) ганглиев. В контексте представления о сердце как о сложной авторегуляторной системе чувствительная иннервация сердца (как вегетативная, так и соматическая) должна рассматриваться как часть системы регуляции
кровотока.
адренергические рецепторы плазмолеммы разных клеток сердца (кардиомиоциты рабочие и особенно атипические, внутрисердечные нейроны собственного нервного аппарата). Существует множество фармакологических препаратов, оказывающих непосредственное действие на названные рецепторы. Так, норадреналин, адреналин и другие адренергические препараты в зависимости от эффекта на а- и p-адренорецепторы подразделяют на активирующие (адреномиметики) и блокирующие (адреноблока- торы) агенты. м-Холинорецепторы также имеют аналогичные классы препаратов (холиномиметики и холиноблокаторы).
а. Активация симпатических нервов увеличивает частоту спонтанной деполяризации мембран водителей ритма, облегчает проведение импульса в волокнах Пуркинье и увеличивает частоту и силу сокращения типичных кардиомиоцитов.
б. Парасимпатические влияния, наоборот, уменьшают частоту генерации импульсов пейсмейкерами, снижают скорость проведения импульса в волокнах Пуркинье и уменьшают частоту сокращения рабочих кардиомиоцитов.
а. Спинальная. Периферические отростки чувствительных нейронов спинномозговых узлов образуют свободные и инкапсулированные нервные окончания.
б. Специализированные сенсорные структуры сердечно-сосудистой системы рассмотрены в главе 10.
Г. Регенерация. При ишемической болезни сердца (ИБС), атеросклерозе коронарных сосудов, сердечной недостаточности разной этиологии (в т.ч. при артериальной гипертензии, инфаркте миокарда) наблюдаются патологические изменения кардиомиоцитов, включая их гибель.
а. Причины сердечной недостаточности — снижение сократительной способности, увеличение посленагрузки, изменения преднагрузки.
Снижение сократительной способности
(а) Инфаркт миокарда — некроз участка сердечной мышцы с потерей его способности к сокращению. Замещение поражённой части стенки желудочков соединительной тканью приводит к снижению функциональных свойств миокарда. При поражении значительной части миокарда развивается сердечная недостаточность.
(б) Врождённые и приобретённые пороки сердца приводят к перегрузке полостей сердца давлением или объёмом с развитием сердечной недостаточности.
(в) Артериальная гипертензия. Многие больные гипертонической болезнью или симптоматическими гипертензиями страдают недостаточностью кровообращения. Снижение сократительной способности миокарда характерно для стойкой тяжёлой гипертензии, быстро приводящей к развитию сердечной недостаточности.
(г) Кардиомиопатии токсические (алкоголь, кобальт, катехоламины, доксору- бицин), инфекционные, при т.н. коллагеновых болезнях, рестриктивные (ами- лоидоз и саркоидоз, идиопатические).
б. Компенсаторные механизмы при сердечной недостаточности. Феномены, вытекающие из закона Франка-Старлинга, в т.ч. гипертрофия миокарда, дилатация левого желудочка, периферическая вазоконстрикция вследствие выброса катехоламинов, активация системы ренин-ангиотензин-[альдостерон] и вазопрессина, перепрограммирование синтеза миозинов в кардиомиоцитах, увеличение секреции атриопептина, — компенсаторные механизмы, поддерживающие положительный инотропный эффект. Однако рано или поздно миокард теряет способность обеспечивать нормальный сердечный выброс.