Поперечно-полосатая сердечная мышечная ткань
Поперечно-полосатая сердечная мышечная ткань
Лекция по гистологии №10
Мышечные ткани.
Мышечные ткани — это ткани для которых способность к сокращению является главным свойством.
Классификация мышечной ткани.
По строению
По происхождению
o Мезенхимная – гладкая мышечная ткань стенок полых органов и ресничного тела глаз млекопитающих
o Эпидермальная – гладка мышечная ткань концевых отделов и выводных протоков желез развиващихся из кожной эктодермы
o Нейральная – развивается из глазных бокалов, являющихся выростами нервной трубки, к данной группе относятся мышцы, суживающие и расширяющие зрачок
o Целомическая – сердечная мышечная ткань
o Миотомная (соматическая) – к данной группе относится скелетная поперечно-полосатая мышечная ткань
Строение поперечно-полосатой мышечной ткани:
Основной элемент ткани: истинное мышечное волокно, которая фактически является симпластом (многоядерная структура, все ядра находятся под одной плазмолеммой). Длина мышечного волокна соответствует длине мышцы, диаметр 20-50мкм. Снаружи мышечное волокно покрыто сарколеммой (мясо). Сарколемма состоит из двух мембран:
Между этими мембранами располагаются миосателитоциты – это мелкие клетки с одним округлым ядром выполняющие роль камбия. Ядра мышечных волокон располагаются под плазмолеммой, имеют вытянутую палочковидную форму. Количество до нескольких тысяч и в следствии отсутствия в миосимпластах центриолей ядра не способны к делению. Цитоплазма мышечного волокна – саркоплазма, в ней находятся органоиды общего значения из которых наиболее хорошо развита гладкая ЭПС и митохондрии, имеются включения гликогена, липидов и миоглобина – это белок способный связывать кислород имеющий в своем составе атомы железа. Хорошо развиты миофибриллы, которые являются сократительным аппаратом волокна, располагаются вдоль его оси, за их счет волокно имеет поперечную исчерченость. Миофибриллы состоят из миофиломентов: тонких актиновых и толстых миозиновых. Филаменты располагаются упорядоченно и их концы находятся на одинаковом уровне. При этом концы актиновых филоментов заходят между концами миозиновых филаментов. Пространственно вокруг одного миозинового филамента находится 6 концов актиновых филоментов. Участок миофибриллы образованный только актиновыми филаментами называетсяИ-диском (изотропный или светлый). Через центр этого диска проходит Z-линия (телофрагма) –она состоит из альфа-актинина, к ней прикрепляются актиновые филаменты. Миозиновые филаменты вместе с заходящими между ними концами актиновых филаментов образуют диск А (амизатропный или темный или диск с двойным лучепреломлением). Диск А разделяется М-линией (мезофрагма) –состоит из белканиомиозина. В центредиска Аимеется H-полоскаограниченная концами актиновых филаментов. Структурно-функциональной единицей ниофибрилл является Саркомер.
Саркомер –участок миофибрилл расположенный между двумя телофрагмами или Z-линиями.
Сокращение мышечного волокна возможно только после поступления к нему нервного импульса. Для передачи возбуждения от плазмолеммы к миофибриллам существуют мембранные структуры к ним отнстся:
o Т-трубочки – глубокие каналообразные впячивания плазмалеммы идущие в поперечном направлении вокруг миофибрил.
o L-канальцы – являются компонентами гладкой эндо-плазматический сети, которая располагается петлеобразно вдоль каждой миофибриллы.
В области Т-трубочек L-канальцы расширяются, образуя конечные или терминальные цистерны, которые окружают Т-трубочки с двух сторон образуя триады.
В L-канальцыах находятся ионы кальция без которых невозможно сокращение, так как центры взаимодействия милофеломентов заблокированы белком тропомиозином.Ионы кальция осуществляют разблокирование после чего концы актиновых филоментов вдвигаются между миозиновыми филаментами. При этом концы миазиновых филаментов
Приближаются к телофрагмы концы актиновых филментов приближаются к мезофрагмам, уменьшается размер диска-И и размер H-полоски, после прекращения потенциала дествия идущего по Т-трубочкам ионы кальция возвращаются в L-канальцы, центры взаимодействия миофиламентов блокируются белком происходит расслобление мышечного волокна.
Строение мышцы как органа:
Каждое мышечное волокно окружено тонкой прослойкой рыхлой волокнистой соединительной ткани, которая называется эндомизий. В эндомизии проходят кровеносные и лимфатические сосуды и нервные окончания. Несколько мышечных волокон объединяются в пучок, который окружен перемизием.Вся мышца окруженаэпимизием.
Мион –мышечное волокно вместе с сосудами и нервами.
Нервно-мышечная единица –группа мышечных волокон инервируемая одним мото-нейроном.
Мышечные пуски образуют мясистую часть мышцы, которая называется брюшком. Брюшко переходит в сухожилие которое необходимо для прикрепления мышцы к скелету. Это происходит так: на концах мышечных волокон имеются впячивания сарколеммы, в которые входят коллагеновые волокна сухожилий, проникают через базальную мембрану и при помощи молекулярных соединений связываются с плазмолеммой как бы привязывая коллагеновое волокно к мышечному.
Два основных типа мышечных волокон:
1 тип – красные или медленные волокна – характеризируются большим количеством миоглобина, чем обусловлен цвет волокон, большим количеством липидных включений, низкой скоростью распада АТФ, малым содержание гликогена и высокой активностью фермента сукценатдегидрогеназы, который необходим для окислительного распада большинства субстратов, данные волокна обладают малой утомляемостью, но длительной сокращаемостью, это так называемый стаерский или статический тип деятельности
2тип – белые, быстрые – они обладают противоположными свойствами, быстрой сокращаемостью и быстрой утомляемостью, это спринтерский динамический тип деятельности.
Развитие мышечной ткани:
Развивается из мезодермальных сомитов, при этом клетки миотомов могут диференцироваться в двух направлениях: из одних будут формироваться – миосателитоциты – необходимые для роста мышечных волокон в детском возрасте и при незначительных повреждениях мышцы; из других клеток миотомов формируются миобласты – в процессе эмбрионального развития сливаются в мышечные трубочки, по мере накопления миофибриллы оттесняют ядра на периферию формируя зрелые мышечные волокна.
Регенерация: Процесс регенерации возможен двумя путями:
1-восстановление целостности мышечного волокна за счет образования мышечных почек, при этом на концах разорванных волокон гипертрофируется гранулярная ЭПС, которая начинает синтезировать белки волокна, в результате концы мышечных волокон утолщаются и приближаются друг к другу, но между ними успевает прорасти соединительная тканьэндомизияформируя рубец.
2-образование новых мышечных волокон – возможно за счет дифференцировки миосателитоцитов и превращение их в миобласты.
Поперечно-полосатая сердечная мышечная ткань
Развивается из миоэпикардиальной пластинки, висцеральных листков спланхнатомов и образует среднюю оболочку сердца – миокард. Она состоит из: типичных кардиомиоцитов (сократительные), атипичных кардиомиоцитов (проводящих), секреторных кардиомиоцитов (эндокринные).
Типичные кардиомиоциты – имеют цилиндрическую форму, содержит 1-2 ядра, которые из-за относительно небольшого количества миофибрилл не оттесняются от периферии а находятся в центре. В саркоплазме хорошо развиты гладкая ЭПС, митохондрии, имеются включения гликогена, липидов и миоглобина.
Строение миофибрилл и сокращение мышечной клетки аналогичны предыдущей ткани.
Типичные кардиомиоциты – объединяются в функциональные мышечные волокна. Места соединения кардиомиоцитов называются вставочными дисками, здесь имеются контакты трех типов:
Интердигитации, Десмосомы, Нексусы, через которые происходит обмен веществами и электрическая связь. Функциональные волокна покрыты сарколеммой и связаны друг с другом анастомозами (боковыми соединениями), диаметр функциональных волокон составляет 10-20 мкм.
Атипичные кардиомиоцитами – характеризуются большими размерами до 50 мкм, более светлой цитоплазмой, центрально или эксцентрично расположенным ядром, малым содержанием и неупорядоченным расположение миофибрилл, отсутствием Т-канальцев и более простым устройством вставочных дисков. Формируют проводящую систему сердца.
Функция: выработка и передача сигнала на типичные кардиомиоциты.
Секреторные кардиомиоциты – имеют небольшие размеры, звездчатую форму и располагаются только в предсердиях в своей цитоплазме содержат гранулы секрета, в гранулах которых находятся натриуретический фактор, который регулирует сократимость сердечной мышцы, объем циркулирующей крови, артериальное давление и диурез.
Регенерация —Возможна только внутриклеточная путем гипертрофии оставшихся клеток, волокна данной ткани не восстанавливаются, а замещаются соединительной тканью.
Гладкая мышечная ткань
Состоит из гладких миоцитов, которые имеют веретенообразную форму длиной до 500мкм, диаметром 6-8мкм снаружи покрыты сарколеммой, плотно прилежат к друг другу соединяются при помощи нексусов. Ядра имеют палочковидную форму, располагаются в центре клетки, в цитоплазме находятся слабо развитые органеллы общего значения, но из них хорошо развиты гладкая и гранулярная ЭПС и митохондрии располагающиеся у полюсов ядра, лизосомы, комплекс Гольджи и клеточный центр, также имеются включения гликогена, липидов и миоглобина, сократительным аппаратом являются миофиламенты актиновые. Актиновые филаменты располагающиеся вдоль клетки либо под углом длинной оси, в местах соединения актиновых филаментов друг с другом и с плазмолеммой имеются плотные тельца, являющиеся аналогами телофрагм. Миозиновые филаменты появляются только при поступлении в клетки нервного импульса, так как в покои они диссацированны на фрагменты и отдельные молекулы (поэтому отсутствуют миофибриллы и не будет поперечных исчерченностей).
В гладких миоцитах нет Т-трубочек и L-канальцев, поэтому под влияние нервного импульса, концентрация ионов кальция повышается за счет открытия кальциевых каналов и за счет кавеол.
Кавеолы – это пиноцитозные пузырьки содержащие ионы кальция и образующиеся из многочисленных впячиваний плазмалеммы. Ионы кальция запускают сократительный процесс, при этом концы актиновых филаментов также сдвигаются между концами миазиновых филаментов. Сила тяги прилагается к потным тельцам и к плазмолемме, что вызывает сокращение миоцита.
Регенерация — возможна путем гипертрофии – то есть увеличение объема оставшихся клеток и путем гиперплазии – увеличение числа гладких миоцитов за счет их митотического деления или путем дифференцировки миофибробластов.
http://lektsii.org/6-88550.html
Мышцы, органы тела животных и человека, за счет сокращения и расслабления которых происходят все движения тела и внутренних органов. Мышцы образуются мышечной тканью в сочетании с другими тканевыми структурами — соединительнотканными компонентами, нервами и кровеносными сосудами.
Мышечная ткань состоит из мышечных клеток, которым в наибольшей степени присуще свойство сократимости. Издавна эти клетки называют мышечными волокнами; данный термин указывает лишь на то, что мышечные клетки имеют вытянутую форму. Сократимость (одно из основных свойств животных клеток, которое наиболее развито в мышечных тканях) мышечных волокон обеспечивается аппаратом, образованным сократительными белками (актином и миозином), взаимодействие которых, протекающее с использованием энергии (АТФ), приводит к сокращению клеток (укорочению). Вслед за сокращением наступает расслабление, и тогда они возвращаются к своей исходной длине. Благодаря такому свойству клеток мышечной ткани достигается все многообразие двигательных функций организма и протекающих в нем механических процессов.
Существует три типа мышечной ткани и соответственно мышц, различающихся по структуре мышечных волокон и характеру иннервации:
1. Скелетная (поперечно-полосатая) мышечная ткань
2. Сердечная поперечно — полосатой мышечная ткань
3. Гладкая мышечная ткань
Скелетная (поперечно-полосатая) мышечная ткань — упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов; один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания.
Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) диаметром от 50 до 100 мкм. Клетки многоядерные, содержат до 100 и более ядер. Микроскопическое изучение показало, что волокно скелетной мышцы по всей длине имеет регулярную поперечную исчерченность в виде чередующихся светлых и темных участков, (поперечнополосатая мышечная ткань образована мышечными клетками, содержащими миофибриллы, которые состоят из миозиновых и актиновых протофибрилл, взаимное положение которых создает поперечную исчерченность),что послужило основанием для другого названия — поперечнополосатые мышцы.
Функции скелетных мышц находятся под контролем центральной нервной системы, т.е. контролируются нашей волей, поэтому их называют также произвольными мышцами. Однако они могут находиться в состоянии частичного сокращения и независимо от нашего сознания; такое состояние называют тонусом. мышечный ткань волокно
Сердечная поперечно — полосатая мышечная ткань
Структурно-функциональной единицей сердечной поперечно — полосатой мышечной ткани является клетка — кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
— типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
— атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50-120 мкм в длину, шириной 15-20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть — функциональный синтиций. Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (дисков) между кардиомиоцитами нет. Вставочные диски — это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы — аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов — атипичные кардиомиоциты образуют проводящую систему сердца, состоящую из:
— предсердно-желудочковый пучок (пучок Гиса), ствол, правую и левую ножки;
— концевые разветвления ножек — волокна Пуркинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичных рядом особенностей:
— они крупнее (длина 100 мкм, толщина 50 мкм);
— в цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
— плазмолемма не образует Т-канальцев;
— во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
· Р-клетки (пейсмекеры) — водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов — миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнее из целомического эпителия этих участков.
http://studbooks.net/1281875/meditsina/serdechnaya_poperechno_polosataya_myshechnaya_tkan
Поперечно-полосатая мышечная ткань
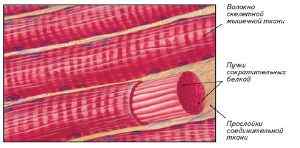

Поперечно-полосатая мышечная (ПМ) ( англ. musculi transversostriati ) — сократимая ткань, состоящая из симпластов — многоядерных мышечных волокон, покрытых возбудимой плазматической мембраной — сарколеммой, сходной по электрическим свойствам с мембранами нервных клеток.
Группы волокон образуют мышечные пучки, которые, объединяясь, образуют мышцу. В соединительной ткани, окружающей мышечные волокна, мышечные пучки и всю мышцу, проходят кровеносные сосуды и нервы. ПМ имеют видимую в световой микроскоп поперечную исчерченность (отсюда название), обусловленную чередованием в миофибриллах участков с разными физико-химическими и оптическими свойствами. ПМ составляют у позвоночных скелетную (туловищную, или соматическую) мускулатуру. В зависимости от соотношения в волокнах ПМ количества саркоплазмы и миофибрилл различают белые ПМ, которые содержащат относительно мало саркоплазмы и много миофибрилл, способные сильно сокращаться, но быстро утомляющиеся, и красные ПМ, богатые саркоплазмой и относительно бедные миофибриллами, сокращающиеся с меньшей силой, но способные к длительной работе. По сравнению с белыми ПМ саркоплазма красных содержит гораздо больше митохондрий и миоглобина. В целом ПМ сокращаются быстрее, чем гладкие мышцы. Нервные окончания в ПМ сигнализируют в ЦНС о состоянии мышечной ткани, а из ЦНС в мышечные волокна поступают нервные импульсы, вызывающие их возбуждение и сокращение. К ПМ относят и сердечную мышцу (миокард), основу которой составляют отдельные клетки — кардиомиоциты. Из беспозвоночных ПМ имеются у некоторых моллюсков, например, головоногих моллюсков — осьминогов, кальмаров и других имеются поперечнополосатые мышцы, что связано с их подвижным образом жизни. Особенно сильно развиты они в ноге животного. ПМ обеспечивают в основном функцию перемещения тела или отдельных его частей в пространстве. (см. МЫШЦЫ). [1]
Поперечно-полосатая скелетная мышечная ткань Править
Клетки поперечно-полосатой скелетной мышечной ткани имеют большую длину (до нескольких см) и диаметр 50—100 мкм; эти клетки многоядерные, содержат до 100 и более ядер; в световом микроскопе цитоплазма выглядит как чередование тёмных и светлых полосок. Свойствами этой мышечной ткани является высокая скорость сокращения, расслабления и произвольность (то есть её деятельность управляется по воле человека). Эта мышечная ткань входит в состав скелетных мышц, а также стенки глотки, верхней части пищевода, ею образован язык, глазодвигательные мышцы. Волокна длиной от 10 до 12 см.
Поперечно-полосатая сердечная мышечная ткань Править
Поперечно-полосатая сердечная мышечная ткань состоит из одно- или двухъядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы (по периферии цитолеммы). Кардиомиоциты разветвлены и образуют между собой соединения — вставочные диски, в которых объединяется их цитоплазма. Существует также другой межклеточный контакт — анастомозы (впячивание цитолеммы одной клетки в цитолемму другой). Этот вид мышечной ткани образует миокард сердца. Развивается из миоэпикардальной пластинки (висцерального листка спланхнотома шеи зародыша). Особым свойством этой ткани является автоматия — способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках (типичные кардиомиоциты). Эта ткань является непроизвольной (атипичные кардиомиоциты). Существует третий вид кардиомиоцитов — секреторные кардиомиоциты (в них нет фибрилл). Они синтезируют предсердный натрийуретический пептид (атриопептин) — гормон, вызывающий снижение объёма циркулирующей крови и системного артериального давления. [2]
Сердечная поперечно-полосатая мышечная ткань;
Структурно-функциональной единицей является клетка —кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
· типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
· атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50—120 мкм в длину, шириной 15—20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть — функциональный синтиций. Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (диском между кардиомиоцитами нет. Вставочные диски — это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы — аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов — атипичные кардиомиоциты образуют проводящую систему сердца, состоящую из:
· предсердно-желудочковый пучок (пучок Гиса)ствол, правую и левую ножки;
· концевые разветвления ножек — волокна Пункинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичным рядом особенностей:
· они крупнее (длина 100 мкм, толщина 50 мкм);
· в цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
· плазмолемма не образует Т-канальцев;
· во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
· Р-клетки (пейсмекеры) водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов — миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнееиз целомического эпителия этих участков.
Иннервация сердечной мышечной ткани
Биопотенциалы сократительные кардиомиоциты получают из двух источников:
· из проводящей системы сердца (прежде всего из синусо-предсердного узла);
· из вегетативной нервной системы (из ее симпатической и парасимпатической части).
Регенерация сердечной мышечной ткани
Кардиомиоциты регенерируют только по внутриклеточному типу. Пролиферации кардиомиоцитов не наблюдается. Камбиальные элементы в сердечной мышечной ткани отсутствуют. При поражении значительных участков миокарда (в частности, при инфаркте миокарда) восстановление дефекта происходит за счет разрастания соединительной ткани и образования рубцов (пластическая регенерация). Естественно, что сократительная функция в этих участках отсутствует. Поражение проводящей системы сопровождается нарушением ритма сердечных сокращений.
http://studopedia.su/6_8755_serdechnaya-poperechno-polosataya-mishechnaya-tkan.html
Классификация сократимых тканей.
Мышечными тканями (textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма (сердце, язык, кишечник и др.).
Свойством изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
Основные морфологические признаки элементов мышечных тканей: удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина. Специальные сократительные органеллы — миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков — актина и миозина — при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин — белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко падает).
Классификация мышечных тканей:
Гладкая мышечная ткань внутренних органов
Поперечно-полосатая мышечная ткань скелетной мускулатуры
Поперечно-полосатая мышечная ткань сердца
Гистогенетическая (по Н.Г. Хлопину)
Мезенхимные мышечные ткани
Эпидермальные мышечные ткани
Нейральные мышечные ткани
Мезодермальные мышечные ткани

Поперечно-полосатая сердечная мышечная ткань
Сердечная мышечная ткань, которая по строению и функции отличается от скелетных мышц, состоит из кардиомиоцитов, образующих соединяющиеся друг с другом комплексы. По своему строению сердечная мышечная ткань похожа на скелетную (поперечнополосатая исчерченность), однако сокращения сердечной мышцы не подконтрольны сознанию человека, она иннервируется вегетативной нервной системой.
Строение миофибрилл аналогично таковому скелетных мышц. Однако, в отличие от последних, между мио-фибриллами кардиомиоцитов нет столь четких границ. По периферии клетки и между митохондриями находится множество частичек гликогена и элементов гладкого эндоплазматического ретикулума. В кардиомиоцитах имеется очень большое количество крупных митохондрий с хорошо развитыми кристами, которые располагаются группами между миофибриллами. На уровне Z-линий плазмалемма кардиомиоцитов также формирует Т-трубочки, вблизи которых сосредоточены скопления цистерн гладкого эндоплазматического ретикулума. Однако триады выражены менее четко, чем в скелетных мышцах. Кардиомиоциты соединены между собой вставочными дисками . На участках вставочного диска, лежащих параллельно продольной оси кардиомиоцита , находятся лентовидные десмосомы (пояски сцепления, к ним, возможно, прикрепляются актиновые филаменты) и щелевидные контакты , не связанные с миофиламентами. Через нексусы (щелевидные контакты) осуществляются передача нервного возбуждения и обмен ионами между клетками.
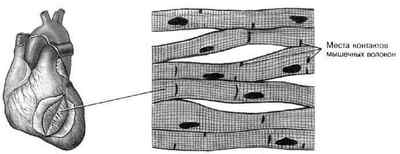
Поперечно-полосатая мышечная ткань скелетной мускулатуры
Скелетная (поперечно-полосатая) мышечная ткань — упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов: один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 70-75 % из воды.
Источником развития скелетной мускулатуры являются клетки миотомов — миобласты. Часть из них дифференцируется в местах образования так называемых аутохтонных мышц. Прочие же мигрируют из миотомов в мезенхиму; при этом они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникает 2 клеточные линии. Клетки первой сливаются, образуя симпласты — мышечные трубки (миотубы). Клетки второй группы остаются самостоятельными и дифференцируются в миосателлиты (миосателлитоциты).
В первой группе происходит дифференцировка специфических органелл миофибрилл, постепенно они занимают большую часть просвета миотубы, оттесняя ядра клеток к периферии.
Клетки второй группы остаются самостоятельными и располагаются на поверхности мышечных трубок.
Структурной единицей мышечной ткани является мышечное волокно. Оно состоит из миосимпласта и миосателлитоцитов (клеток-спутниц), покрытых общей базальной мембраной.
Длина мышечного волокна может достигать нескольких сантиметров при толщине в 50-100 микрометров.

Данный вид мышечной ткани обеспечивает возможность выполнения произвольных движений. Сокращающаяся мышца воздействует на кости или кожу, к которым она прикрепляется. При этом один из пунктов прикрепления остаётся неподвижным — так называемая точка фиксации, которая в большинстве случаев рассматривается в качестве начального участка мышцы. Перемещающийся фрагмент мышцы называют подвижной точкой, которая является местом её прикрепления.
http://studfiles.net/preview/5811238/
Химия, Биология, подготовка к ГИА и ЕГЭ
В биологии высших животных различают 4 вида тканей. Одна из них — мышечная.
За счет мышц обеспечивается не только передвижение организма в пространстве, но и работа внутренних органов.
Мы рассмотрим виды этой ткани, особенности клеточного строения и характерные функции.
Мышечная ткань

Клетки мышечной ткани
- Форма клетки — вытянутая;
- в состав входит миофибрилл и миофиламенты — органеллы, за счет которых клетки обладают главным свойством мышечной ткани — сократимостью;
- в клетках больше митохондрий (у них больше затраты энергии);
- они могут быть многоядерными.
Гладкая мышечная ткань
- Клетки содержат одно ядро;
- Это ткань внутренних органов — легких, желудочно-кишечного тракта, кровеносных и лимфатических сосудов. Сокращается и расслабляется медленно, автоматически , и, соответственно, не произвольно.
Этим видом мышечной ткани человек не управляет, она отвечает за работу внутренних органов.
Еще один вид ткани внутреннего органа — одного из самых важных в организме — сердца.
Сердечная поперечно-полосатая мышечная ткань

Сердце — орган, который должен работать бесперебойно. Следовательно, ткань, составляющая этот орган, должна потреблять много энергии, и быстро регенерировать.
Мышечная ткань сердца млекопитающих не имеет возможности восстанавливаться после повреждений (исключение — млекопитающие в эмбриональном периоде способны к регенерации органа в определённых рамках), в отличие от тканей некоторых рыб и амфибий. Однако исследователи Юго-Западного медицинского центра системы Техасского университета доказали, что сердце мышонка, которому всего день от рождения ещё может восстанавливаться, а сердце семидневного мышонка — уже нет.
- Клетки могут быть одно- или двухъядерные; содержат много крупных митохондрий;
- эти клетки имеют на концах ответвления — отростки — вставочные диски , и таким образом соединяются между собой, образуя сплошную сеть.
Этот вид мышечной ткани присущ только одному органу — сердцу, естественно, работа автоматическая и ткань не утомляется ?
Поперечно-полосатая мышечная ткань
Её еще называют скелетной. Этот вид ткани отвечает именно за передвижение тела животного в пространстве.
- Клетки многоядерные, при чем количество ядер может достигать 100;
- Обеспечивает быстрые движения, при этом быстро утомляется;
- Работа этой мышечной ткан и не автоматическая.
- в ЕГЭ это вопросы А15, А16 и А17
- A33 — Процессы жизнедеятельности организма человека
- A34 — Человек. Нейрогуморальная регуляция
- часть С — вопросы по анатомии
- в ГИА — А9 — Анатомия и физиология человека
http://distant-lessons.ru/myshechnaya-tkan.html
Мышечная система человека
- Физиология
- История физиологии
Общий обзор мышечной системы человека
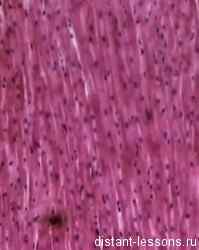
У позвоночных животных и человека различают три разных по строению группы мышц:
- поперечно-полосатые мышцы скелета;
- поперечно-полосатая мышца сердца;
- гладкие мышцы внутренних органов, сосудов и кожи.
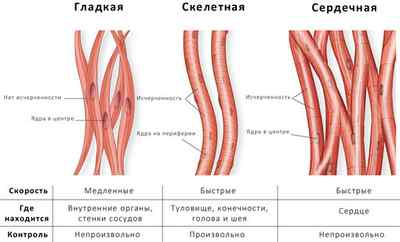
Рис. 1. Виды мышц человека
Гладкие мышцы
Из двух видов мышечной ткани (поперечно-полосатой и гладкой) гладкая мышечная ткань находится на более низкой ступени развития и присуща низшим животным.
Гладкие мышцы образуют мышечный слой стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других полых органов. Они состоят из веретенообразных мышечных волокон и не имеют поперечной исчерченности, так как миофибриллы в них расположены менее упорядоченно. В гладких мышцах отдельные клетки соединяются между собой специальными участками наружных мембран — нексусами. За счет этих контактов потенциалы действия распространяются с одного мышечного волокна на другое. Поэтому в реакцию возбуждения быстро вовлекается вся мышца.
Гладкие мышцы осуществляют движения внутренних органов, кровеносных и лимфатических сосудов. В стенках внутренних органов они, как правило, располагаются в виде двух слоев: внутреннего кольцевого и наружного продольного. В стенках артерии они формируют спиралевидные структуры.
Характерной особенностью гладких мышц является их способность к спонтанной автоматической деятельности (мышцы желудка, кишечника, желчного пузыря, мочеточников). Это свойство регулируется нервными окончаниями. Гладкие мышцы пластичны, т.е. способны сохранять приданную растяжением длину без изменения напряжения. Скелетная мышца, наоборот, обладает малой пластичностью и эту разницу легко установить в следующем опыте: если растянуть с помощью грузов и гладкую и поперечно-полосатую мышцы и снять груз, то скелетная мышца сразу же после этого укорачивается до первоначальной длины, а гладкая мышца долгое время может находиться в растянутом состоянии.
Такое свойство гладких мышц имеет большое значение для функционирования внутренних органов. Именно пластичность гладких мышц обеспечивает лишь небольшое изменение давления внутри мочевого пузыря при его наполнении.
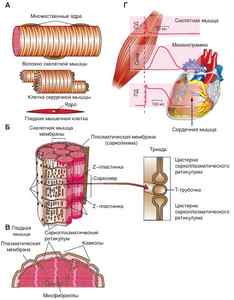
Рис. 2. А. Волокно скелетной мышцы, клетка сердечной мышцы, гладкая мышечная клетка. Б. Саркомер скелетной мышцы. В. Строение гладкой мышцы. Г. Механограмма скелетной мышцы и мышцы сердца.
Гладким мышцам присущи те же основные свойства, что и поперечнополосатым скелетным мышцам, но и некоторые особые свойства:
- автоматия, т.е. способность сокращаться и расслабляться без внешних раздражений, а за счет возбуждений, возникающих в них самих;
- высокая чувствительность к химическим раздражителям;
- выраженная пластичность;
- сокращение в ответ на быстрое растяжение.
Сокращение и расслабление гладких мышц происходит медленно. Это способствует наступлению перестальтических и маятникообразных движений органов пищеварительного тракта, что приводит к перемещению пищевого комка. Длительное сокращение гладких мышц необходимо в сфинктерах полых органов и препятствует выходу содержимого: желчи в желчном пузыре, мочи в мочевом пузыре. Сокращение гладкомышечных волокон совершается независимо от нашего желания, под воздействием внутренних, не подчиненных сознанию причин.
Поперечно-полосатые мышцы
Поперечно-полосатые мышцы располагаются на костях скелета и сокращением приводят в движение отдельные суставы и все тело. Скелетные мышцы образуют тело, или сому, поэтому их еще называют соматическими, а иннервирующую их систему — соматической нервной системой.
Благодаря деятельности скелетной мускулатуры осуществляется передвижение тела в пространстве, разнообразная работа конечностей, расширение грудной клетки при дыхании, движение головы и позвоночника, жевание, мимика лица. Насчитывается более 400 мышц. Общая масса мышц составляет 40% веса. Обычно средняя часть мышцы состоит из мышечной ткани и образует брюшко. Концы мышц — сухожилия построены из плотной соединительной ткани; они соединяются с костями при помощи надкостницы, но могут прикрепляться и к другой мышце, и к соединительному слою кожи. В мышце мышечные и сухожильные волокна объединяются в пучки при помощи рыхлой соединительной ткани. Между пучками располагаются нервы и кровеносные сосуды. Сила мышцы пропорциональна количеству волокон, составляющих брюшко мышцы.
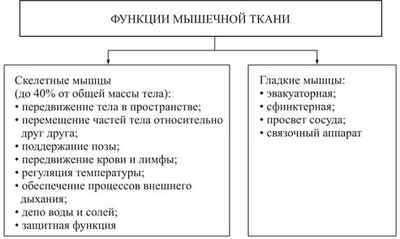
Рис. 3. Функции мышечной ткани
Некоторые мышцы проходят только через один сустав и при сокращении приводят его в движение — односуставные мышцы. Другие мышцы проходят через два или несколько суставов — многосуставные, они производят движение в нескольких суставах.
При сокращении концы мышцы, прикрепленные к костям, приближаются друг к другу, а размеры мышцы (длина) уменьшается. Кости, соединенные суставами, действуют как рычаги.
Изменяя положение костных рычагов, мышцы действуют на суставы. При этом каждая мышца влияет на сустав только в одном направлении. У одноосного сустава (цилиндрический, блоковидный) имеются две действующие на него мышцы или группы мышц, являющиеся антагонистами: одна мышца — сгибатель, другая — разгибатель. В то же время на каждый сустав в одном направлении действует, как правило, две мышцы и более, являющиеся синергистами (синергизм — совместное действие).
У двуосного сустава (эллипсоидный, мышелковый, седловидный) мышцы группируются соответственно двум его осям, вокруг которых совершаются движения. К шаровидному суставу, имеющему три оси движения (многоосный сустав), мышцы прилежат со всех сторон. Так, например, в плечевом суставе имеются мышцы-сгибатели и разгибатели (движения вокруг фронтальной оси), отводящие и приводящие (сагиттальная ось) и вращатели вокруг продольной оси, кнутри и кнаружи. Различают три вида работы мышц: преодолевающую, уступающую и удерживающую.
Если благодаря сокращению мышцы меняется положение части тела, то преодолевается сила сопротивления, т.е. выполняется преодолевающая работа. Работа, при которой сила мышцы уступает действию силы тяжести и удерживаемого груза, называется уступающей. В этом случае мышца функционирует, однако она не укорачивается, а удлиняется, например, когда невозможно поднять или удержать на весу тело, имеющее большую массу. При большом усилии мышц приходится опустить это тело на какую-нибудь поверхность.
Удерживающая работа выполняется благодаря сокращению мышц, тело или груз удерживается в определенном положении без перемещения в пространстве, например человек держит груз, не двигаясь. При этом мышцы сокращаются без изменения длины. Сила сокращения мышц уравновешивает массу тела и груза.
Когда мышца, сокращаясь, перемешает тело или его части в пространстве, они выполняют преодолевающую или уступающую работу, которая является динамической. Статистической является удерживающая работа, при которой не происходит движений всего тела или его части. Режим, при котором мышца может свободно укорачиваться, называется изотоническим (не происходит изменения напряжения мышцы и меняется только ее длина). Режим, при котором мышца не может укоротиться, называется изометрическим — меняется только напряжение мышечных волокон.
Рис. 4. Мышцы человека
Строение поперечно-полосатых мышц
Скелетные мышцы состоят из большого числа мышечных волокон, которые объединяются в мышечные пучки.
В одном пучке содержится 20-60 волокон. Мышечные волокна представляют собой клетки цилиндрической формы длиной 10-12 см и диаметром 10-100 мкм.
Каждое мышечное волокно имеет оболочку (сарколемму) и цитоплазму (саркоплазму). В саркоплазме находятся все компоненты животной клетки и вдоль оси мышечного волокна располагаются тонкие нити — миофибриллы, Каждая миофибрилла состоит из протофибрилл, в состав которых вкючены нити белков миозина и актина, являющихся сократительным аппаратом мышечного волокна. Миофибриллы разделены между собой перегородками, которые называются Z-мембранами, на участки — саркомеры. На обоих концах саркомеров к Z-мембране прикреплены тонкие актиновые нити, а в середине расположены толстые миозиновые нити. Нити актина своими концами частично входят между миозиновыми нитями. В световом микроскопе нити миозина выглядят в виде светлой полоски в темном диске. При электронной микроскопии скелетные мышцы выглядят исчерченными (поперечно-полосатыми).
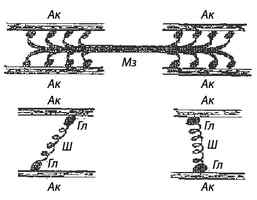
Рис. 5. Поперечные мостики: Ак — актин; Мз — миозин; Гл — головка; Ш — шейка
На боковых сторонах миозиновой нити имеются выступы, получившие название поперечных мостиков (рис. 5), которые расположены под углом 120° по отношению к оси миозиновой нити. Актиновые филаменты выглядят в виде двойной нити, закрученной в двойную спираль. В продольных бороздках актиновой спирали находятся нити белка тропомиозина, к которым присоединен белок тропонин. В состоянии покоя молекулы белка тропомиозина расположены таким образом, чтобы предотвращать прикрепление поперечных мостиков миозина к актиновым нитям.
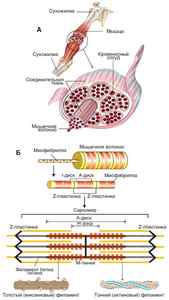
Рис. 6. А — организация цилиндрических волокон в скелетной мышце, прикрепленной к костям сухожилиями. Б — структурная организация филаментов в волокне скелетной мышцы, создающая картину поперечных полос.
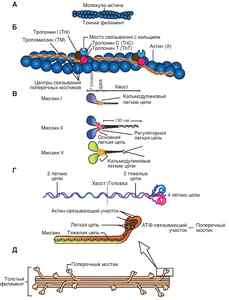
Рис. 7. Строение актина и миозина
Во многих местах поверхностная мембрана углубляется в виде микротрубок внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-система). Параллельно миофибриллам и перпендикулярно поперечным трубочкам между миофибрилл расположена система продольных трубочек (саркоплазматический ретикулум). Концевые расширения этих трубочек — терминальные цистерны — подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В цистернах сосредоточено основное количество внутриклеточного кальция.
Механизм сокращения скелетной мышцы
Мышечная ткань состоит из клеток, называемых мышечными волокнами. Снаружи волокно окружено оболочкой — сарколеммой. Внутри сарколеммы содержится цитоплазма (саркоплазма), содержащая ядра и митохондрии. В ней содержится огромное количество сократительных элементов, называемых миофибриллами. Миофибриллы проходят от одного конца мышечного волокна до другого. Они существуют сравнительно короткий срок — около 30 суток, после чего и происходит их полная смена. В мышцах идет интенсивный синтез белка, необходимый для образования новых миофибрилл.
Мышечное волокно содержит большое количество ядер, которые располагаются непосредственно под сарколеммой, поскольку основная часть мышечного волокна занята миофибриллами. Именно наличие большого числа ядер обеспечивает синтез новых миофибрилл. Такая быстрая смена миофибрилл обеспечивает высокую надежность физиологических функций мышечной ткани.
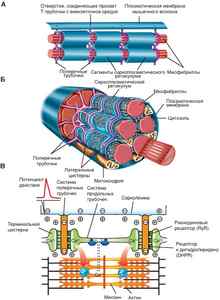
Рис. 7. А — схема организации саркоплазматического ретикулума, поперечных трубочек и миофибрилл. Б — схема анатомической структуры поперечных трубочек и саркоплазматического ретикулума в индивидуальном волокне скелетной мышцы. В — роль саркоплазматического ретикулума в механизме сокращения скелетной мышцы
Каждая миофибрилла состоит из правильно чередующихся светлых и темных участков. Эти участки, обладая разными оптическими свойствами, создают поперечную исчерченность мышечной ткани.
В скелетной мышце сокращение вызывается поступлением к ней импульса по нерву. Передача нервного импульса с нерва на мышцу осуществляется через нервно-мышечный синапс (контакт).
Одиночный нервный импульс, или однократное раздражение, приводит к элементарному сократительному акту — одиночному сокращению. Начало сокращения не совпадает с моментом нанесения раздражения, поскольку существует скрытый, или латентный, период (интервал между нанесением раздражения и началом сокращения мышцы). В этот период происходит развитие потенциала действия, активация ферментных процессов и распад АТФ. После этого начинается сокращение. Распад АТФ в мышце приводит к превращению химической энергии в механическую. Энергетические процессы всегда сопровождаются выделением тепла и тепловая энергия обычно является промежуточной между химической и механическими энергиями. В мышце же химическая энергия превращается непосредственно в механическую. Но тепло в мышце образуется и за счет укорочения мышцы, и во время ее расслабления. Тепло, образующееся в мышцах, играет большую роль в поддержании температуры тела.
В отличие от сердечной мышцы, которая обладает свойством автоматики, т.е. она способна сокращаться под влиянием импульсов, возникающих в ней самой, и в отличие от гладкой мускулатуры, также способной к сокращению без поступления сигналов извне, скелетная мышца сокращается только при поступлении к ней сигналов из ЦНС. Непосредственно сигналы к мышечным волокнам поступают по аксонам двигательных клеток, расположенным в передних рогах серого вещества спинного мозга (мотонейронам).
Рефлекторный характер деятельности мышц и координация мышечных сокращений
Скелетные мышцы в отличие от гладких способны совершать произвольные быстрые сокращения и производить этим значительную работу. Рабочим элементом мышцы является мышечное волокно. Типичное мышечное волокно представляет собой структуры с несколькими ядрами, отодвинутыми на периферию массой сократительных миофибрилл.
Мышечные волокна обладают тремя основными свойствами:
- возбудимостью — способностью отвечать на действия раздражителя генерацией потенциала действия;
- проводимостью — способностью проводить волну возбуждения вдоль всего волокна в обе стороны от точки раздражения;
- сократимостью — способностью сокращаться или изменять напряжение при возбуждении.
В физиологии имеется понятие двигательной единицы, под которой подразумевается один двигательный нейрон и все мышечные волокна, которые этот нейрон иннервирует. Двигательные единицы бывают разными по объему: от 10 мышечных волокон на единицу для мышц, выполняющих точные движения, до 1000 и более волокон на двигательную единицу для мышц «силовой направленности». Характер работы скелетных мышц может быть различным: статическая работа (поддержание позы, удержание груза) и динамическая работа (перемещение тела или груза в пространстве). Мышцы участвуют также в передвижении крови и лимфы в организме, выработке тепла, актах вдоха и выдоха, являются своеобразными депо для воды и солей, защищают внутренние органы, например мышцы брюшной стенки.
Для скелетной мышцы характерны два основных режима сокращения — изометрический и изотонический.
Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, при попытке поднять очень большой груз), — она не укорачивается.
Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способное поднять данный груз, а потом мышца укорачивается — меняет свою длину, сохраняя напряжение, равное весу удерживаемого груза. Чисто изометрического или изотонического сокращения практически наблюдать нельзя, но существуют приемы так называемой изометрической гимнастики, когда спортсмен напрягает мышцы без изменения длины. Эти упражнения в большей мере развивают силу мышц, чем упражнения с изотоническими элементами.
Сократительный аппарат скелетной мышцы представлен миофибриллами. Каждая миофибрилла диаметром 1 мкм состоит из нескольких тысяч протофибрилл — тонких, удлиненных полимеризированных молекул белков миозина и актина. Миозиновые нити в два раза тоньше актиновых, и в состоянии покоя мышечного волокна актиновые нити свободными кольцами входят между миозиновыми нитями.
В передаче возбуждения большую роль играют ионы кальция, которые входят в межфибриллярное пространство и запускают механизм сокращения: взаимное втягивание относительно друг друга актиновых и миозиновых нитей. Втягивание нитей происходит при обязательном участии АТФ. В активных центрах, расположенных на одном из концов миозиновых нитей, АТФ расщепляется. Энергия, выделяемая при расщеплении АТФ, преобразуется в движение. В скелетных мышцах запас АТФ невелик — всего на 10 одиночных сокращений. Поэтому необходим постоянный ре- синтез АТФ, который идет тремя путями: первый — за счет запасов креатинфосфата, которые ограничены; второй — гликолитический путь при анаэробном расщеплении глюкозы, когда на одну молекулу глюкозы образуется две молекулы АТФ, но одновременно образуется молочная кислота, которая тормозит активность гликолитических ферментов, и наконец третий — аэробное окисление глюкозы и жирных кислот в цикле Кребса, совершающееся в митохондриях и образующее 38 молекул АТФ на 1 молекулу глюкозы. Последний процесс наиболее экономичный, но очень медленный. Постоянная тренировка активизирует третий путь окисления, в результате чего повышается выносливость мышц к длительным нагрузкам.
http://www.grandars.ru/college/medicina/myshcy-cheloveka.html