СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ — Основы физиологии
СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ
Способность сердца сокращаться в течение всей жизни без остановки обусловлена рядом специфических физических и физиологических свойств сердечной мышцы.
Физические свойства. Растяжимость — способность увеличивать длину без нарушения структуры под влиянием растягивающей силы. Такой силой является кровь, наполняющая полости сердца во время диастолы. От степени растяжения мышечных волокон сердца в диастолу зависит сила их сокращения в систолу.
Эластичность — способность восстанавливать исходное положение после прекращения действия деформирующей силы. Эластичность сердечной мышцы является полной, т.е. она полностью восстанавливает исходные показатели.
Способность развивать силу в процессе сокращения мышцы.
Физиологические свойства. Сокращения сердца происходят вследствие периодически возникающих процессов возбуждения в сердечной мышце, которая обладает рядом физиологических свойств: автоматизмом, возбудимостью, проводимостью, сократимостью.
Способность сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом, носит название автоматизм.
В сердце различают сократительную мускулатуру, представленную поперечно-полосатой мышцей, и атипическую, или специальную ткань, в которой возникает и проводится возбуждение. Атипическая мышечная ткань содержит малое количество миофибрилл, много саркоплазмы и не способна к сокращению. Она представлена скоплениями в определенных участках миокарда, которые образуют проводящую систему сердца, состоящую из синоатриального узла, располагающегося на задней стенке правого предсердия у места впадения полых вен; атриовентрикулярного, или предсердно-желудочкового узла, находящегося в правом предсердии вблизи перегородки между предсердиями и желудочками; предсердно-желудочкового пучка (пучка Гиса), отходящего от атриовентрикулярного узла одним стволом. Пучок Гиса, пройдя через перегородку между предсердиями и желудочками, разветвляется на две ножки, идущие к правому и левому желудочкам. Заканчивается пучок Гиса в толще мышц волокнами Пуркинье.
Синоатриальный узел является водителем ритма первого порядка. В нем возникают импульсы, которые определяют частоту сокращений сердца. Он генерирует импульсы со средней частотой 70—80 импульсов в 1 мин.
Атриовентрикулярный узел — водитель ритма второго порядка.
Пучок Гиса — водитель ритма третьего порядка.
Волокна Пуркинье — водители ритма четвертого порядка. Частота возбуждения, возникающая в клетках волокон Пуркинье, очень низкая.
В норме атриовентрикулярный узел и пучок Гиса являются только передатчиками возбуждений из ведущего узла к сердечной мышце.
Однако и они обладают автоматизмом, только в меньшей степени, и этот автоматизм проявляется лишь при патологии.
В области синоатриального узла обнаружено значительное число нервных клеток, нервных волокон и их окончаний, которые образуют здесь нервную сеть. К узлам атипической ткани подходят нервные волокна от блуждающих и симпатических нервов.
Возбудимость сердечной мышцы — способность клеток миокарда при действии раздражителя приходить в состояние возбуждения, при котором изменяются их свойства и возникает потенциал действия, а затем сокращение. Сердечная мышца менее возбудима, чем скелетная. Для возникновения возбуждения в ней необходим более сильный раздражитель, чем для скелетной. При этом величина реакции сердечной мышцы не зависит от силы наносимых раздражений (электрических, механических, химических и др.). Сердечная мышца максимально сокращается и на пороговое, и на более сильное по величине раздражение.
Уровень возбудимости сердечной мышцы в разные периоды сокращения миокарда меняется. Так, дополнительное раздражение сердечной мышцы в фазу ее сокращения (систолу) не вызывает нового сокращения даже при действии сверхпорогового раздражителя. В этот период сердечная мышца находится в фазе абсолютной рефрактерности. В конце систолы и начале диастолы возбудимость восстанавливается до исходного уровня — это фаза относительной рефрактерности. За этой фазой следует фаза экзальтации, после которой возбудимость сердечной мышцы окончательно возвращается к исходному уровню. Таким образом, особенностью возбудимости сердечной мышцы является длительный период рефрактерности.
Проводимость сердца — способность сердечной мышцы проводить возбуждение, возникшее в каком-либо участке сердечной мышцы, к другим ее участкам. Возникнув в синоатриальном узле, возбуждение распространяется по проводящей системе на сократительный миокард. Распространение этого возбуждения обусловлено низким электрическим сопротивлением нексусов. Кроме того, проводимости способствуют специальные волокна.
Волны возбуждения проводятся по волокнам сердечной мышцы и атипической ткани сердца с неодинаковой скоростью. Возбуждение по волокнам мышц предсердий распространяется со скоростью 0,8—1 м/с, по волокнам мышц желудочков — 0,8—0,9 м/с, по атипической ткани сердца — 2—4 м/с. При прохождении возбуждения через атриовентрикулярный узел возбуждение задерживается на 0,02- 0,04 с — это атриовентрикулярная задержка, обеспечивающая координацию сокращения предсердий и желудочков.
Сократимость сердца — способность мышечных волокон укорачиваться или изменять свое напряжение. Она реагирует на раздражители нарастающей силы по закону «все или ничего». Сердечная мышца сокращается по типу одиночного сокращения, так как длительная фаза рефрактерности препятствует возникновению тетани- ческих сокращений. В одиночном сокращении сердечной мышцы выделяют: латентный период, фазу укорочения (систола), фазу расслабления (диастола). Благодаря способности сердечной мышцы сокращаться только по типу одиночного сокращения сердце выполняет функцию насоса.
Первыми сокращаются мышцы предсердий, затем слой мышц желудочков, обеспечивая тем самым движение крови из полостей желудочков в аорту и легочный ствол.
http://studref.com/348067/agropromyshlennost/svoystva_serdechnoy_myshtsy
Физиология сердца Свойства сердечной мышцы
Сердечная мышца обладает следующими свойствами: 1)автоматией —способностью сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом; 2)возбудимостью —способностью сердца приходить в состояние возбуждения под действием раздражителя; 3)проводимостью —способностью сердечной мышцы проводить возбуждение; 4)сократимостью —способностью изменять свою форму и величину под действием раздражителя, а также растягивающей силы или крови.
Субстратом автоматии в сердце является специфическая щечная ткань, илипроводящая система сердца,которая состоит изсинусно-предсердного(синоатриального)(СА) узла,расположенного в стенке правого предсердия у места впадения в него верхней полой вены,предсердно-желудочкового(атриовентрикулярного)узла,расположенного в межпредсердной перегородке на границе предсердий и желудочков. От атриовентрикулярного узла начинается пучокГиса.Пройдя в толщу межжелудочковой перегородки, он делится на правую и левую ножки, заканчивающиеся конечными разветвлениями —волокнами Пуркинье.Верхушка сердца не обладает автоматией, а лишь сократимостью, так как в ней отсутствуют элементы проводящей системы сердца.
В нормальных условиях водителем ритма, или пейсмекером, является синоатриальный узел. Частота разрядов синоатриального узла в покое составляет 70в 1минуту. Атриовентрикулярный узел —это водитель ритма второго порядка с частотой 40 — 50в 1 минуту. Он берет на себя роль водителя ритма, если по каким-либо причинам возбуждение от СА не может перейти на предсердия при атриовентрикулярной блокаде или при нарушении проводящей системы желудочков. Если поражены все основные водители ритма, то очень редкие импульсы (20имп/с) могут возникать в волокнах Пуркинье —это водитель ритма 3-го порядка.
Следовательно, существует градиент автоматии сердца,согласно которому степень автоматии тем выше, чем ближе расположен данный участок проводящей системы к синусному узлу.
Электрическая активность клеток миокарда и проводящей системы сердца
Потенциал действия кардиомиоцитов начинается с быстрой риверсии мембранного потенциала, составляющего -90мВ и создаваемого за счет К + -потенциала, до пика ПД (+30мВ) (рис.11). Этофаза быстрой деполяризации,обусловленная коротким значительным повышением проницаемости дляNa\’ 1 \», который лавинообразно устремляется в клетку. Фаза быстрой деполяризации очень короткая и составляет всего 1—2мс. Начальный входNа + быстро инактивируется, однако деполяризация мембраны продолжается за счет активации медленных натрий-кальциевых каналов, а вход Са 2+ приводит к развитиюплато ПД —это специфическая особенность клеток миокарда. В этот период быстрые натриевые каналы инактивируются и клетка становится абсолютно невозбудима. Этофаза абсолютной рефрактерности.Одновременно происходит активация калиевых каналов, а выходящие из клетки ионы К + создаютфазу быстрой реполяризации мембраны.
Ускорение процесса реполяризации происходит за счет закрытия кальциевых каналов. В конце периода реполяризации постепенно закрываются калиевые каналы и реактивируются натриевые. Это приводит к восстановлению возбудимости кардиомиоцита и возникновению относительной рефракторной фазы. Длительность ПД кардиомиоцита составляет 200 — 400мс.
Р


ис. 11. Схемы потенциалов действия различных отделов сердца, кривой сокращения и фаз возбудимости сердечной мышцы: А —схема потенциала действия клетки миокарда (/),кривой сокращения (II)и фаз возбудимости (III) сердечной мышцы; 1 —потенциал действия клетки миокарда: / —быстрая деполяризация; 2 —пик, 3 —плато, 4 —быстрая ре поляризация;II —кривая сокращения: а —фаза сокращения,б —фаза расслабления;III —кривая возбудимости: 5 —абсолютная рефракторная фаза, б —относительная рефракторная фаза, 7 —фаза супернормальной возбудимости;Б —схема потенциала действия клетки пейсмекера (синоаурикулярного узла): МДП —максимальный диастолический потенциал; МДД —медленная диастолическая деполяризация
Калий-натриевый насос, создающий потенциал покоя или мембранный потенциал миокардиоцита, может быть инактивирован под действием сердечных гликозидов (препараты наперстянки, строфантина), которые приводят также к повышению внутриклеточной концентрации Na + , снижению интенсивности обмена внутриклеточногоCa 2+ на внеклеточныйNа + , накоплению Ca 2+ в клетке. В результате сократимость миокарда становится больше. Ее можно увеличить и за счет повышения внеклеточной концентрации Са 2+ и с помощью веществ (адреналин, норадреналин),ускоряющих вход Са 2+ во время ПД. Если удалить Са 2+ из внешней среды или заблокировать вход Ca 2+ во время ПД с помощью таких веществ —антагонистов кальция, как верапамил, нифедипин и др., то сократимость сердца уменьшается.
Клетки проводящей системы сердца и, в частности, клетки пейсмекера, обладающие автоматией, в отличие от клеток рабочего миокарда-кардиомиоцитов могут спонтанно деполяризоваться до критического уровня. В таких клетках за фазой реполяризации следует фаза медленной диастолической деполяризации. (МДД),которая приводит к снижению МП до порогового уровня и возникновению ПД. МДД —это местное, нераспространяющееся возбуждение, в отличие от ПД, который является
Таким образом, пейсмекерные клетки отличаются от кардиомиоцитов: 1)низким уровнем МП —около 50—70мВ, 2)наличием МДД, 3)близкой к пикообразному потенциалу формой ПД,4)низкой амплитудой ПД — 30-50мВ без явления риверсии (овершута).
Особенности электрической активности пейсмекерных клеток обусловлены целым рядом процессов, происходящих на их мембране. Во-первых, эти клетки даже в условиях «покоя» имеют повышенную проницаемость для ионов Na + , что приводит к снижению МП. Во-вторых, в период реполяризации на мембране открываются только медленные натрий-кальциевые каналы, так как быстрые натриевые каналы из-за низкого МП уже инактивирова-ны. В клетках синоатриального узла в период реполяризации быстро инактивируются открытые калиевые каналы, но повышается натриевая проницаемость, на фоне которой и возникает МДД, а затем и ПД. Потенциал действия синоатриального узла распространяется на все остальные отделы проводящей системы сердца.
Таким образом, синоатриальный узел навязывает всем «ведомым» отделам проводящей системы свой ритм. Если возбуждение не поступает от главного пейсмекера, то «латентные» водители ритма, т.е. клетки сердца, обладающие автоматией, берут на себя функцию нового пейсмекера, в них также зарождается МДД и ПД, а сердце продолжает свою работу.
http://studfiles.net/preview/3549695/
Средства, влияющие на возбудимость, проводимость сердечной мышцы и ритм сердечных сокращений
Средства, влияющие на возбудимость, проводимость сердечной мышцы и ритм сердечных сокращений
Сердечный ритм зависит от автоматии, возбудимости и проводимости сердечной мышцы. Аритмии – нарушения ритма деятельности сердца. Они могут возникать вследствие повышения или угнетения автоматии в синоатриальном узле, а также в результате повышения автоматии или в эктопическом, или в латентном водителе ритма. Увеличение автоматии синоатриального узла называется синусовой тахикардией, уменьшение – синусовой брадикардией.
Одной из причин возникновения аритмий может быть так называемый «механизм повторного входа возбуждения» (reentrумеханизм), который имеет место при пароксизмальной тахикардии, мерцании и трепетании предсердий и желудочков, экстрасистолии.
Нарушение проводимости – наиболее частая причина возникновения аритмий, включает в себя замедление или блокаду проведения импульсов, о которых говорилось выше.
Для лечения аритмий используют антиаритмические препараты, которые по механизму действия делят на пять групп.
К первой группе относят препараты с мембраностабилизирующим действием (уменьшают проницаемость клеточной мембраны и нарушают транспорт К + , Na + и Са 2+ , блокируют ацетилхолин). Эти вещества угнетают автоматию клеток синоатриального узла и эктопических водителей ритма, снижают возбудимость волокон Пуркинье и миофибрилл, уменьшают скорость проведения импульсов через атриовентрикулярный узел и волокна Пуркинье, увеличивают продолжительность ПД и абсолютной рефрактерной фазы. К этой группе препаратов относят хинидин, новокаинамид, ритмодан и др. Их принимают при мерцательной аритмии, трепетании предсердий, пароксизмальной тахикардии, частой предсердной и желудочковой экстрасистолии, желудочковой тахикардии.
Препараты К + – панангин, аспаркам, хлорид калия приводят к снижению мембранного потенциала миоцитов и уменьшению скорости проведения импульсов в миофибриллах. Их назначают при аритмиях, связанных с гипокалиемией, и при аритмиях, вызванных передозировкой дигиталиса.
Ко второй группе антиаритмических препаратов относят лидокаин, мекситил, пропафенон и др. В отличие от хинидина, эти препараты уменьшают рефрактерный период, повышают скорость проведения импульсов, не влияя на сократимость миокарда. Кроме того, они обладают местноанестезирующим действием и показаны при остром инфаркте миокарда. Их используют для лечения желудочковой экстрасистолии и профилактики фибрилляции желудочков сердца.
Антиаритмические препараты третьей группы – это блокаторы бета-адренергических рецепторов. Они уменьшают автоматик) предсердий и желудочков, снижают атриовентрикулярную и внутрижелудочковую проводимость, укорачивают время реполяризации. Их назначают при желудочковой экстрасистолии, трепетании и мерцании предсердий. К препаратам этой группы относят обзидан, оксипренолол, аптин, вискен.
Антиаритмические препараты четвертой группы обладают антиаритмическими свойствами, не влияя на мембраны за счет блокады постганглионарной симпатической передачи импульсов. Они укорачивают ПД и абсолютный рефрактерный период. Представителем этой группы является препарат кордарон. Его назначают при пароксизмальных аритмиях, синусной тахикардии, трепетании предсердий, желудочковой тахикардии и экстрасистолии.
К пятой группе антиаритмических препаратов относят антагонисты кальция, например верапамил (изоптин, финоптин). Он тормозит трансмембранный ток Са 2+ внутри сердечной клетки, уменьшает спонтанную активность синоатриального узла.
http://med.wikireading.ru/24952
Проводимость сердечной мышцы
Сокращение сердца, как и у скелетных мышц, запускается ПД. Однако если у скелетной мышцы ПД составляет всего несколько миллисекунд и предшествует сокращению, то у сердечной ПД и фазы сокращения перекрывают друг друга. ПД заканчивается только после начала фазы расслабления. Это одна из особенностей электромеханического сопряжения сердечной мышцы. Другая особенность состоит в том, что существует взаимосвязь между внутриклеточным депо Са 2+ и Са 2+ внеклеточной среды. Как упоминалось выше, во время ПД Са 2+ входит в клетку из внеклеточной среды и увеличивает длительность ПД, а значит, и рефракторного периода, тем самым создаются условия для пополнения внутриклеточных запасов кальция, участвующего в последующих сокращениях сердца.
Электрокардиография
Вокруг возбужденного сердца возникает электрическое поле, которое можно зарегистрировать с поверхности тела в виде электрокардиограммы. Электрические потенциалы прежде всего возникают в возбужденном синоатриальном узле. Этот участок становится электроотрицательным по отношению к невозбужденному, заряженному положительно. Это и приводит к появлению электрических потенциалов и дальнейшему их распространению по проводящей системе сердца, миокарду предсердий и желудочков.
Электрокардиограмма отражает процесс возникновения возбуждения и его проведение по сердцу, но не его сокращение. В нормальной электрокардиограмме различают пять зубцов: Р, Q, R, S, Т(рис.12). Возникновение зубцаРобусловлено распространением возбуждения в предсердиях —это алгебраическая сумма электрических потенциалов, возникающих в предсердиях. ЗубецQсоответствует возбуждению сосочковых мышц. Зубец R —возбуждению оснований желудочков, зубец S —верхушки сердца. Зубец Г отражает процесс реполяризации желудочков и состояние метаболизма миокарда. Он очень изменчив и может искажаться при различного рода интоксикациях, например, при инфекциях (дизентерия и др.), отравлениях химическими ядами, при гипоксии, инфаркте миокарда, диабете.
Итак, различают предсердный комплекс, куда входит зубец Р, исегментPQ,а также желудочковый комплексQRSи сегментST. Интервал PQот начала зубцаРдо начала зубца Qотражает время проведения возбуждения от предсердий к желудочкам, в норме он равен 0,12-0,18с.
При нарушении проведения импульсов из предсердий к желудочкам, вызванном или органическими изменениями в проводящей системе, или отравлением сердечными глюкозидами, увеличением содержания ионов К + , снижением МП, а также гипоксией возникаетнеполная атриовентрикулярная блокада.При этом не все импульсы периодически проводятся к желудочкам или их проведение задерживается, тогда интервалPC? становится больше 0,18с.
При полном нарушении проводимости между предсердиями и желудочками возникает полная атриовентрикулярная блокада —предсердия и желудочки сокращаются независимо друг от друга: предсердия в синусном ритме, желудочки —в ритме пейсмекера 2-го или 3-го порядка.
Р

ис. 12. Электрокардиограмма (схема зубцов и интервалов) ЗубецРотражает возбуждение предсердий; интервалP—Q —распространение возбуждений от предсердий к желудочкам;Q—T (QRST) —желудочковый комплекс; зубец Т —процесс реполяризации желудочков
Длительность комплекса ORSсоставляет 0,06 — 0,1с. Его уши-рение является признаком нарушения внутрижелудочковой проводимости. ИнтервалQTсоставляет 0,36с и зависит от частоты сердечных сокращений. Чем больше частота, тем короче интервал. Амплитуда зубцов ЭКГ следующая: Р 0,6[/i]MB; T=от 1/6до2/3R.
Для регистрации ЭКГ используют 3стандартных биполярных отведения от конечностей (треугольник Эйнтховена), 1-е отведение: правая рука-левая рука; 2-е отведение: правая рука-левая нога; 3-е отведение: левая рука-левая нога. Кроме того, регистрируют 3усиленных униполярных отведения:aVR —активный электрод на правой руке,aVL —активный электрод на левой руке,aVF —активный электрод на левой ноге и 6униполярных грудных отведении по Вильсону —V1—V6
При биполярных отведениях по Эйнтховену точки, от которых отводят потенциалы, совпадают с вершинами равностороннего треугольника, стороны которого и представляют собой оси отведении. С помощью треугольника Эйнтховена можно установить величину электродвижущей силы сердца, а значит, и высоту зубцов ЭКГ. Высота зубца Rво 2-м отведении в нормограмме равна сумме зубца Rв 1-ми 3-м отведении, т.е.R2 =R1+R3
http://studfiles.net/preview/6667095/page:3/
Возбудимость и проводимость сердца
- Физиология
- История физиологии
- Методы физиологии
Особенности возбудимости, проводимости и сократимости сердечной мышцы
Сократительные кардиомиоциты представлены клетками длиной около 100-150 мкм и толщиной около 10-15 мкм (миокард желудочков), 40-70 мкм и 5-6 мкм соответственно (миокард предсердий). Они имеют многие общие черты строения и обладают функциями, присущими другим типам клеток, в частности клеткам скелетных мышц. В то же время кардиомиоциты характеризуются рядом особенностей строения и свойств, обеспечивающих выполнение ими функций возбуждения, проведения возбуждения и сокращения, специфических для миокарда.
Возбудимость сердечной мышцы
Важнейшими особенностями возбудимости сердечной мышцы являются:
- способность возбуждаться в ответ на поступление потенциала действия из проводящей системы сердца;
- способность более длительно находиться в состоянии возбуждения, по сравнению с волокнами скелетной мышцы;
- наличие длительного периода абсолютной рефрактерности, почти совпадающего по времени с длительностью одиночного сокращения (систолы).
Возбудимость обусловлена свойствами плазматической мембраны (сарколеммы) сократительного кардиомиоцита, в которой имеются разнообразные ионные каналы. Мембрана состоит из внутреннего слоя толщиной около 10 нм и наружного примембранного слоя гликокаликса. Основой клеточной мембраны кардиомиоцитов является фосфолипидный бислой, в котором каналообразующие белки формируют натриевые, кальциевые, калиевые и другие типы каналов. На внутренней поверхности мембраны имеются области, с которыми связано большое количество ионов кальция. Этот пул кальциевых ионов может быстро высвобождаться внутрь клетки во время возбуждения или обмениваться с внеклеточным пространством.
В состоянии покоя (в диастолу) трансмембранная разность потенциалов кардиомиоцита составляет около 85-90 мВ (фаза 4-го мембранного потенциала). Это обусловлено разностью концентраций ионов но обе стороны мембраны, высокой плотностью в ней Na+ / К+ АТФазы и примерно в 10 раз более высокой проницаемостью мембраны в покос для ионов K + , чем в миоцитах скелетных мышц. Соотношение проницаемостей в мембране сократительного кардиомиоцита Рк+: PNa+: РCI-= = 1 : 0,05: 0,1.
Поступающий из проводящей системы к кардиомиоцитам через каналы щелевых контактов потенциал действия деполяризует их мембрану. Когда величина деполяризации достигает критического уровня (около -60 мВ), изменяют свою пространственную конформацию активационные ворота быстрых потенциалзависимых натриевых каналов и через открывающиеся каналы поток ионов Na + по градиентам электрического поля и концентрации устремляется в миоцит, быстро деполяризует и вскоре перезаряжает мембрану — фаза 0 потенциала действия (рис. 1). Проницаемость мембраны для ионов Na+ за период времени, пока открыты активационные ворота, возрастает в несколько сот раз но сравнению с проницаемостью в состоянии покоя. Смена заряда на мембране, поляризованной до +20 мВ, сопровождается закрытием инактивационных ворот натриевых каналов и прекращением входа ионов Na+ в миоцит. Инактивационные ворота натриевых каналов остаются закрытыми до момента, пока мембрана клетки не будет частично реполяризована в фазу 3 потенциала действия. В течение времени от момента активации — открытия натриевых каналов в фазу 0 до их реактивации в фазу 3 мембрана кардиомиоцитов находится в состоянии абсолютной рефрактерности и, таким образом, никакие воздействия на ее в это время не могут вызвать на мембране новый процесс возбуждения и сокращения миокарда.
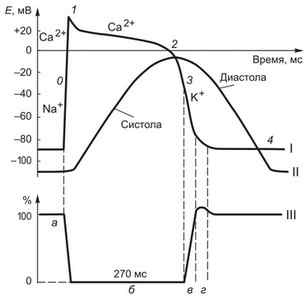
Рис. 1. Соотношение во времени процессов возбуждения, возбудимости и сокращения в сократительных миоцитах миокарда: а — исходная возбудимость; б — период абсолютной невозбудимости (рефрактерности); в — период относительной рсфрактсрности; г — период повышенной возбудимости; I — потенциал действия сократительных кардномиоцнтов (0, 1, 2, 3, 4 — фазы потенциала действия); II — сокращение миокарда; III изменение возбудимости миокарда в процессе возбуждения
Во время деполяризации мембраны наряду с активацией натриевых каналов происходит активация и открытие нескольких типов медленных потенциалзависимых калиевых каналов. Выход ионов К+ из клетки через эти каналы по концентрационному градиенту, но против градиента электрического ноля, запаздывает но отношению ко входу в клетку ионов Na + . Суммарный ноток выходящих ионов К+ начинает существенно превышать вход ионов Na+ с момента инактивации проницаемости натриевых каналов. Когда мембрана клетки приобретает изнутри избыток положительных зарядов, ионы К+ выходят из клетки уже не только но градиенту их концентраций, но и по градиенту электрического поля. Выход ионов К+ обусловливает развитие фазы быстрой реполяризации мембраны миоцита (фаза 1 потенциала действия).
Деполяризация мембраны кардиомиоцита до уровня около -70 мВ сопровождается открытием потенциалзависимых кальциевых каналов Т-типа, через которые в клетку поступает некоторое количество ионов Са 2+ . Однако плотность этих каналов в мембране кардиомиоцитов невелика. При деполяризации мембраны до уровня около -10 мВ активируются медленные потенциалзависимые кальциевые каналы L-типа. Вскоре после начала реполяризации мембраны миоцита вход положительных зарядов ионов Са 2 + по градиенту концентрации в клетку сравнивается с выходом положительных зарядов ионов К+ из клетки, скорость реполяризации замедляется, а фаза 1 потенциала сменяется фазой 2, или фазой плато потенциала действия.
Входящие в фазу плато в кардиомиоцит ионы Са 2 + имеют значение не только для удерживания мембраны в деполяризованном состоянии в течение длительного промежутка времени, но и в высвобождении дополнительного количества кальция из примембранного пула и саркоплазматического ретикулума, а также в сопряжении процессов возбуждения и сокращения миоцитов.
Постепенно развивающаяся инактивация медленных кальциевых каналов ведет к тому, что выход положительно заряженных ионов К через калиевые каналы вновь начинает доминировать над входом ионов Са 2 + и фаза плато сменяется новым ускорением реполяризации мембраны — фазой 3 потенциала. Вскоре мембрана кардиомиоцита реполяризуется до исходного значения трансмембранной разности потенциалов около 90 мВ, характерной для диастолы миокарда (фаза 4).
Таким образом, общая длительность потенциала действия сократительных кардиомиоцитов, вследствие наличия в их мембране ионных каналов медленного типа и других особенностей проницаемости, составляет около 300 мс, что почти в 100 раз превышает длительность потенциала в миоцитах скелетных мышц. Особенно важно, что из 300 мс около 200 мс приходится на фазу плато потенциала действия, во время которой в клетку поступают ионы Ca 2 + , участвующие как в процессе возбуждения, так и в процессе сокращения.
Важное физиологическое значение в регуляции электрических и механических процессов миокарда имеет то, что процессы активации и инактивации потенциалзависимых кальциевых и других ионных каналов мембраны кардиомиоцитов могут модулироваться не только величиной трансмембранной разности потенциалов, но и действием различных сигнальных молекул. Известно, что кальциевые каналы не только потенциалзависимы, но и чувствительны к дигидропиридину. Их проницаемость модулируется при фосфорилировании цАМФ зависимой протеинкиназой А. В настоящее время создан ряд лекарственных веществ (в частности, производных дигидропиридина), с помощью которых оказалось возможным управлять потоками ионов Са 2 + и оказывать влияние на частоту, ритм, силу и другие показатели сокращений сердца.
Восстановление нормального распределения ионов Na+, К+ и Са 2 + по обе стороны мембраны, нарушенного в процессе генерации потенциала действия, осуществляется Na+ /K+ — насосом, кальциевым насосом и натрий кальциевым обменным механизмом.
Состояние возбудимости мембраны сократительного кардиомиоцита в процессе возбуждения определяет возможность возникновения нового процесса возбуждения и последующего сокращения кардиомиоцитов в ходе уже начавшегося возбуждения, т.е. в различные фазы потенциала действия.
В течение времени от начала открытия потенциалзависимых натриевых каналов (фаза 0) и примерно до середины периода конечной реполяризации (-50 мВ), когда становится возможной реактивация инактивированных натриевых каналов (фаза 3), мембрана сократительных кардиомиоцитов находится в состоянии абсолютной невозбудимости (рефрактерности). Продолжительность этого периода составляет около 240 мс при общей продолжительности потенциала действия около 300 мс (рис. 1.3). Состояние абсолютной рефрактерности мембраны кардномиоцитов имеет важное физиологическое значение. Оно заключается в том, что воздействие в этот период на сердце каких-либо стимулов, не оказывает влияния на начавшееся сокращение (систолу) миокарда. Благодаря этому сердечная мышца, в отличие от скелетных мышц, не способна к суммации одиночных сокращений и развитию тетанического сокращения, что предотвращает нарушение насосной функции сердца.
Период абсолютной рефрактерности мембраны сменяется переходом мембраны в состояние относительной рефрактерности. В конце фазы 3 потенциала уже возможно открытие натриевых каналов под действием стимула, сила которого превышает пороговую. В это время на мембране кардиомиоцита способен возникнуть потенциал действия меньшей амплитуды, но он может оказаться достаточным для запуска нового внеочередного сокращения. Период относительной рефрактерности длится около 30 мс.
За несколько десятков миллисекунд до окончания реполяризации мембраны се возбудимость может оказаться более высокой, чем в состоянии покоя. Это объясняется тем, что натриевые каналы к этому времени практически реактивированы, а разность потенциалов близка к уровню Екр. В этот период за короткое мгновение до завершения на мембране предыдущего возбуждения действие даже подпороговых стимулов может вызвать новый процесс возбуждения мембраны (период супернормальной возбудимости).Период повышенной возбудимости кардиологи называют «периодом уязвимости» сердца, так как при поступлении в это время к миокарду внеочередных потенциалов действия возникает повышенная опасность возникновения аритмии, или фибрилляции, желудочков.
Проводимость сердца
Отличается от проводимости скелетной мышцы тем, что в сердце возбуждение может передаваться от одного кардиомиоцита на другие. Таким образом, возникнув в одном участке миокарда, возбуждение может распространяться на его другие участки.
Кардиомиоциты имеют отростчатую форму и контактируют с соседними клетками через нексусы — область плотных контактов между миоцитами, в мембранах которых имеются общие для контактирующих клеток каналы. Каналы сформированы шестью молекулами белков коннексонов и имеют низ- кос сопротивление электрическому току. Они обеспечивают передачу возбуждения с одного кардиомиоцита на другой за счет перехода между клетками ионов и переноса ими электрических зарядов. В частности, через нексусы кардиомиоциты обмениваются ионами Ca 2 + , принимающими участие в передаче возбуждения и сокращения, и другими биологически активными веществами. После того как возбуждение распространилось с одного кардиомиоцита на другой через область вставочных дисков, оно распространяется далее по мембране кардиомиоцитов, благодаря наличию в них потенциалзависимых натриевых, кальциевых, калиевых и других ионных каналов. Скорость проведения возбуждения по миокарду может достигать 1 м/с.
В составе нексусов имеются десмосомы — области прочного механического прикрепления клеток друг к другу. Благодаря наличию непосредственной электрической и механической связи между кардиомиоцитами их возбуждение и сокращение оказываются синхронизированными, а способность миокарда проводить возбуждение и отвечать на него целостной сократительной реакцией получила название функционального синтиция. Поскольку кардиомиоциты способны проводить возбуждение от клетки к клетке, то нет необходимости проведения потенциала действия к каждой клетке волокнами проводящей системы сердца. Между ними и сократительными кардиомиоцитами отсутствуют синапсы.
Таким образом, потенциал действия поступает ко многим точкам миокарда с ветвящихся волокон Пуркинье, где он вызывает возбуждение сократительных кардиомиоцитов, расположенных субэндокардиально. Возникшие в них потенциалы действия распространяются на соседние участки миокарда в направлении от эндокарда к эпикарду. Возникновение возбуждения во многих точках миокарда и его распространение по сердечной мышце с высокой скоростью являются важнейшими условиями синхронизации возбуждения и сокращения не только на уровне отдельных миоцитов, но и в различных участках миокарда. Нарушение распространения возбуждения по миокарду, десинхронизация его сокращения могут быть одной из причин снижения насосной функции сердца.
http://www.grandars.ru/college/medicina/vozbudimost-serdca.html
Возбудимость, проводимость и сократимость сердечной мышцы;
Второй узел расположен в толще сердечной перегородки на границе предсердий и желудочков и называется атриовентрикулярным узлом.
Автоматизм сердечной мышцы
Автоматизм – это способность сердца к ритмическому сокращению под влиянием импульсов, возникающих в самом органе.
Возникновение импульсов связано с функцией особых мышечных клеток – пейсмекеров, которые образуют три узла сердца. Эти три узла обеспечивают выполнение автоматизма сердечной мышцы.
Первый узел расположен в месте впадения полых вен в правое предсердие. Он называется синусным узлом.
Третий узелрасположен ниже второго и носит название пучок Гиса. От пучка Гиса отходят две ножки — правая и левая, каждая из которых заканчивается волокнами Пуркинье, которые образуют синапс с клетками миокарда.
Все три узла связаны между собой межузловыми пучками (которые образованы мышечными клетками) и анатомически образуют единую систему. Данная система проходит через все сердце, от его вершины до основания.
Возникшее возбуждение в первом узле передается по межузловым пучкам ко второму узлу, а затем возбуждение передается на третий узел.
Таким образом, возбуждение охватывает все сердце и вызывает его последовательное сокращение, сначала предсердия, потом желудочки.
Самым главным из трех узлов является первый — синусный узел. Он определяет частоту сердечных сокращений в 1 минуту. Сколько импульсов будет образовано в синусном узле, столько раз сердце сократиться за 1 минуту.
Взаимосвязь между узлами обеспечивает ритмичное сокращение предсердий и желудочков.
В условиях покоя синусный узел вырабатывает 60 – 80 импульсов в минуту, что соответствует 60 – 80 ударам сердца в минуту. Уменьшение ЧСС называется брадикардией, а увеличение – тахикардией.
Возбудимость– это способность сердца отвечать на различные внешние и внутренние раздражения переходом из состояния покоя к состоянию активной деятельности (к сокращению). Возбуждение связано с появлением в возбужденном участке отрицательного электрического потенциала – потенциала действия, который распространяется по сердечной мышце, начиная предсердиями и заканчивая основаниями желудочков. Таким образом, передача возбуждения по сердечной мышце обеспечивается таким свойством, как проводимость.Проводимость наблюдается и при передачи возбуждения от одного узла автоматизма к другому, и к третьему.
Возбудимость сердечной мышцы может изменяться. Если очередное возбуждение застаёт сердечную мышцу в момент сокращения (систолы), то на него сердце не отвечает — это фаза абсолютной рефрактерности(состояние полной невозбудимости). С началом расслабления возбудимость сердца начинает восстанавливаться и наступает фаза относительной рефрактерности. Поступление в этот момент интенсивного стимула вызывает внеочередное сокращение сердца – экстрасистолу. После фазы относительной рефрактерности наступает фаза повышенной возбудимости. Это означает, что в этот момент сокращение сердца могут вызвать даже очень слабые импульсы. Период этот непродолжителен и вскоре наступает восстановление нормального уровня возбудимости.
Возбуждение сердечной мышцы вызывает её сокращение, т.е. увеличение её напряжения или укорочение длины мышечного волокна. Таким образом, сократимость сердца – это способность сердечной мышцы сократиться в ответ на возбуждение
§ 4. Динамика сокращений сердца, фазы сердечного цикла
Безостановочное движение крови по сосудам происходит в результате ритмических сокращений сердца, которые чередуются с его расслаблениями.
Сокращение сердечной мышцы называет систолой, а расслабление – диастолой.
Время, за которое происходит систола предсердий, систола желудочков и общая пауза – называется сердечным циклом.
Во время общей паузы — правое предсердие наполняется кровью из верхней и нижней полой вены, а левое предсердие наполняется кровью из четырех легочных вен. В это время открыты створчатые клапаны и кровь из предсердий течет в желудочки. Полулунные клапаны аорты и легочной артерии — закрыты.
Перед началом систолы желудочков начинается систола предсердий, во время которой происходит некоторое добавочное наполнение желудочков кровью. Однако, это наполнение не играет большой роли, поскольку большая часть наполняющей желудочки крови уже поступила. Сокращение предсердий сменяется диастолой и расслаблением.
После систолы предсердий начинается систола желудочков. Сокращение желудочков развивается постепенно. Вначале увеличивается напряжение мышечной ткани желудочков, что приводит к увеличению в них давления крови. Для того, чтобы кровь из желудочков не вернулась в предсердия, атриовентрикулярные клапаны закрываются. Полулунные клапаны в это время также закрыты, поскольку давление в желудочках ниже, чем в аорте и легочной артерии. По мере увеличения напряжения мышечной ткани желудочков, увеличивается давление крови в них. Когда величина давления в желудочках превысит давление в аорте и легочной артерии, открываются полулунные клапаны и происходит изгнание крови из желудочков.
После изгнания крови из желудочков наступает их диастола.В опустевших желудочках давление становится ниже, чем в сосудах, и для того чтобы кровь не вернулась в сердце закрываются полулунные клапаны. Расслабление желудочков продолжается, давление в них становится еще ниже, и когда величина давления в желудочках станет меньше величины давления в предсердиях откроются створчатые клапаны.
После того, как открываются створчатые клапаны, наступает общая пауза. Кровь из предсердий начинает наполнять желудочки и всё повторяется вновь.
§ 5. Электрокардиограмма (ЭКГ)
Электрокардиограмма – это метод графической регистрации биотоков, возникающих в сердце при его деятельности. По данным ЭКГ можно судить об автоматизме, возбудимости и проводимости сердца.
Для регистрации ЭКГ у человека применяют 3 стандартных отведения. При первом отведении – электроды накладывают на правую и левую руку, при втором – правую руку и левую ногу, при третьем – на левую руку и левую ногу. Помимо стандартных отведений применяют отведения от разных точек грудной клетки, а также однополюсные отведения от конечностей.
Типичная ЭКГ человека состоит из зубцов. Их обозначают латинскими буквами: ?,Q,R,S,T. Три крупных зубца – Р,R,T – обращены вершиной вверх, два мелких – Q, S – направлены вниз. Промежутки между зубцами называют сегментами, совокупность зубца и сегмента – интервалом.
Зубец Р – отражает возбуждение предсердий.
Комплекс Q,R,S – возбуждение желудочков.
Зубец Т – отражает восстановление нормального потенциала мембраны клеток миокарда, т.е. реполяризацию миокарда.
Интервал P-Q отражает время движения возбуждения от предсердий к желудочкам.
Сегмент S-T – соответствует периоду угасания возбуждения желудочков и началу реполяризации.
Интервал Т-Р отражает состояние покоя всей сердечной мышцы (диастола сердца), рисунок 8.
Интервал R-R – отражает длительность сердечного цикла и зависит от частоты сердечных сокращений. При брадикардии он удлинен, при тахикардии – укорочен.
http://studopedia.su/9_72881_vozbudimost-provodimost-i-sokratimost-serdechnoy-mishtsi.html
Автоматия и проводимость сердечной мышцы.
Способность сердца сокращаться в течение всей жизни, не обнаруживая признаков утомления, т.е. автоматизм сердца, связывали вначале с влияниями нервной системы. Однако, постепенно накапливались факты в пользу того, что нейрогенная гипотеза автоматизма сердца, справедливая в отношении многих беспозвоночных животных, не объясняет свойств миокарда у позвоночных. Особенности сокращения сердечной мышцы у последних связывали с функциями атипической ткани миокарда.
В 50-х годах XIX века в опытах Станниуса было показано, что перевязка сердца лягушки на границе между венозным синусом и предсердиями приводит к временной остановке сокращений остальных отделов сердца. Через 30-40 минут сокращения восстанавливаются, однако ритм сокращений области венозного синуса и остальных отделов сердца становится рассогласованным. После наложения второй лигатуры по атриовентикулярной линии прекращается сокращение желудочков с последующим его восстановлением в ритме, не совпадающим, однако, с ритмом сокращений предсердий. Наложение третьей лигатуры в области нижней трети сердца приводит к необратимой остановке сокращений сердца. В дальнейшем было показано, что охлаждение сравнительно небольшого участка в области устья полых вен приводит к остановке сердца.
Результаты этих опытов указывали на то, что в области правого предсердия, а также на границе предсердий и желудочков располагаются участки, ответственные за возбуждение сердечной мышцы. Удалось показать, что сердце человека, извлеченное из трупа и помещенное в теплый физиологический раствор, в результате массажа восстанавливает сократительную активность. Доказано, что автоматизм сердца имеет миогенную природу и обусловлен спонтанной активностью части клеток его атипической ткани. Указанные клетки образуют скопления в определенных участках миокарда. Наиболее важным в функциональном отношении из них является синусный или синоатриальный узел, расположенный между местом впадения верхней полой вены и ушком правого предсердия.
В нижней части межпредсердной перегородки, непосредственно над местом прикрепления септальной створки трехстворчатого клапана, располагается атриовентрикулярный узел. От него отходит пучок атипических мышечных волокон, который пронизывает фиброзную перегородку между предсердиями и переходит в узкий длинный мышечный тяж, заключенный в межжелудочковую перегородку. Он называется атриовентрикулярным пучком или пучком Гиса. Пучок Гиса разветвляется, образуя две ножки, от которых приблизительно на уровне середины перегородки отходят волокна Пуркине, также образованные атипической тканью и формирующие субэндокардиальную сеть в стенках обоих желудочков.
Функция проводимости в сердце имеет электротоническую природу. Она обеспечивается низким электрическим сопротивлением щелевидных контактов (нексусов) между элементами атипического и рабочего миокарда, а также в области вставочных пластинок, разделяющих кардиомиоциты. В результате, сверхпороговое раздражение любого участка вызывает генерализованное возбуждение всего миокарда. Это позволяет считать ткань сердечной мышцы, морфологически разделенную на отдельные клетки, функциональным синцитием.
Возбуждение миокарда зарождается в синоатриальном узле, который называют водителем ритма, или пейсмекером первого порядка, и далее распространяется на мускулатуру предсердий с последующим возбуждением атриовентрикулярного узла, который является водителем ритма второго порядка. Скорость распространения возбуждения в предсердиях составляет в среднем 1 м/с. При переходе возбуждения на атриовентрикулярный узел имеет место так называемая атриовентрикулярная задержка, составляющая 0.04-0.06 с. Природа атриовентрикулярной задержки состоит в том, что проводящие ткани синоатриального и атриовентрикулярного узлов контактируют не непосредственно, а через волокна рабочего миокарда, для которых характерна более низкая скорость проведения возбуждения. Последнее распространяется далее по ножкам пучка Гиса и волокнам Пуркинье, передаваясь на мускулатуру желудочков, которую оно охватывает со скоростью 0.75-4.0 м/с. В силу особенностей расположения волокон Пуркинье, возбуждение сосочковых мышц происходит несколько раньше, чем оно охватывает стенки желудочков. Благодаря этому, нити, удерживающие трехстворчатый и митральный клапаны, оказываются натянутыми раньше, чем на них начинает действовать сила сокращения желудочков. По той же причине наружная часть стенки желудочков у верхушки сердца возбуждается несколько раньше участков стенки, прилежащих к ее основанию.
Указанные сдвиги во времени крайне невелики и обычно принимается, что весь миокард желудочков охватывается возбуждением одновременно. Таким образом, волна возбуждения последовательно охватывает различные отделы сердца в направлении от правого предсердия к верхушке. Данное направление отражает градиент автоматии сердца.
Возбудимость клеток проводящей системы и рабочего миокарда имеет ту же биоэлектрическую природу, что и в поперечнополосатых мышцах. Наличие заряда на мембране здесь также обеспечивается разностью концентраций ионов калия и натрия возле ее внешней и внутренней поверхности и избирательной проницаемостью мембраны для этих ионов. В покое мембрана кардиомиоцитов проницаема для ионов калия и почти непроницаема для натрия. В результате диффузии ионы калия выходят из клетки и создают положительный заряд на ее поверхности. Внутренняя сторона мембраны становится электроотрицательной по отношению к наружной.
В клетках атипического миокарда, обладающих автоматией, мембранный потенциал способен спонтанно уменьшаться до критического уровня, что приводит к генерации потенциала действия. В норме ритм сердечных сокращений задается всего несколькими наиболее возбудимыми клетками синоатриального узла, которые называются истинными водителями ритма или пейсмекерными клетками. В этих клетках во время диастолы мембранный потенциал, достигнув максимального значения, соответствующего величине потенциала покоя (60-70 мВ), начинает постепенно снижаться. Этот процесс называется медленной спонтанной диастолической деполяризацией. Она продолжается до того момента, когда мембранный потенциал достигает критического уровня (40-50 мВ), после чего возникает потенциал действия.
Для потенциала действия пейсмекерных клеток синоатриального узла характерны малая крутизна подъема, отсутствие фазы ранней быстрой реполяризации, а также слабая выраженность «овершута» и фазы «плато». Медленная реполяризация плавно сменяется быстрой. Во время этой фазы мембранный потенциал достигает максимальной величины, после чего вновь возникает фаза медленной спонтанной деполяризации.
Во время диастолы спонтанная деполяризация уменьшает мембранный потенциал (Е мах) до критического уровня (Е кр) и вызывает потенциал действия.
Частота возбуждения пейсмекерных клеток у человека составляет в покое 70- 80 в минуту при амплитуде потенциала действия 70- 80 мВ. Во всех остальных клетках проводящей системы потенциал действия в норме возникает под влиянием возбуждения, приходящего из синоатриального узла. Такие клетки называют латентными водителями ритма. Потенциал действия в них возникает раньше, чем их собственная медленная спонтанная диастолическая деполяризация достигает критического уровня. Латентные водители ритма принимают на себя ведущую функцию только при условии разобщения с синоатриальным узлом. Именно этот эффект наблюдается в упоминавшихся выше опытах Станниуса. Частота спонтанной деполяризации таких клеток у человека составляет 30-40 в минуту.
Спонтанная медленная диастолическая деполяризация обусловлена совокупностью ионных процессов, связанных с функциями плазматических мембран. Среди них ведущую роль играют медленное уменьшение калиевой и повышение натриевой и кальциевой проводимости мембраны во время диастолы, параллельно чему происходит падение активности электрогенного натриевого насоса. К началу диастолы проницаемость мембраны для калия на короткое время повышается, и мембранный потенциал покоя приближается к равновесному калиевому потенциалу, достигая максимального диастолического значения. Затем, проницаемость мембраны для калия уменьшается, что и приводит к медленному снижению мембранного потенциала до критического уровня. Одновременное увеличение проницаемости мембраны для натрия и кальция приводит к поступлению этих ионов в клетку, что также способствует возникновению потенциала действия. Снижение активности электрогенного насоса дополнительно уменьшает выход натрия из клетки и, тем самым, облегчает деполяризацию мембраны и возникновение возбуждения.
Дата добавления: 2015-08-12 ; просмотров: 715 . Нарушение авторских прав