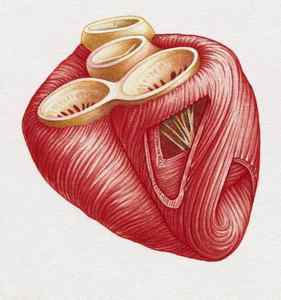
Сердечная поперечно-полосатая мышечная ткань
Сердечная поперечно-полосатая мышечная ткань. Кардиомиоцит. Вставочные диски.
Кардиомиоцит
Структурно-функциональной единицей является клетка —кардиомиоцит.
Классификация кардиомиоцитов
По строению и функциям кардиомиоциты подразделяются на две основные группы:
типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит
представляет собой почти прямоугольную клетку 50—120 мкм в длину, шириной 15—20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой.
В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии.
В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах.
Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть — функциональный синтиций.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (диском между кардиомиоцитами нет.
Вставочные диски
— это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты.
В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты.
Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Атипичные кардиомиоциты
образуют проводящую систему сердца, состоящую из:
предсердно-желудочковый пучок (пучок Гиса)ствол, правую и левую ножки;
концевые разветвления ножек — волокна Пункинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичным рядом особенностей:
они крупнее (длина 100 мкм, толщина 50 мкм);
в цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
плазмолемма не образует Т-канальцев;
во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
Р-клетки (пейсмекеры) водители ритма (I типа);
переходные клетки (II типа);
клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов — миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнееиз целомического эпителия этих участков.
Иннервация сердечной мышечной ткани
Биопотенциалы сократительные кардиомиоциты получают из двух источников:
из проводящей системы сердца (прежде всего из синусо-предсердного узла);
из вегетативной нервной системы (из ее симпатической и парасимпатической части).
Регенерация сердечной мышечной ткани
Кардиомиоциты регенерируют только по внутриклеточному типу. Пролиферации кардиомиоцитов не наблюдается. Камбиальные элементы в сердечной мышечной ткани отсутствуют. При поражении значительных участков миокарда (в частности, при инфаркте миокарда) восстановление дефекта происходит за счет разрастания соединительной ткани и образования рубцов (пластическая регенерация). Естественно, что сократительная функция в этих участках отсутствует. Поражение проводящей системы сопровождается нарушением ритма сердечных сокращений.
Шпаргалка — Электронограммы с пояснениями — файл n1.doc
35. Вставочные диски между кардиомиоцитами (рис. 307)
Вставочный диск между сердечными мышечными клетками миокарда морской свинки. Электронная микрофотограмма. ? 76 000
1 — вставочный диск (граница между мышечными клетками); 2 — сарколемма; 3 — миофибриллы; 4 — митохондрии
Комментарии к электронограмме:
36. Саркомер скелетного мышечного волокна (рис. 158)
Тонкие (актиновые) и толстые (миозиновые) миопротофибриллы. Электронная микрофотограмма поперечно-полосатых миофибрилл. ? 175 000
1 — часть поперечно-полосатой миофибриллы; 2 — толстые (миозиновые) миопротофибриллы (миофиламенты) ; 3 — тонкие (актиновые) миопротофибриллы (миофиламенты); 4 — полоска Т (Z) (телофрагма); 5 — часть I-диска; 6 — полоска М (мезофрагма); 7 — A-диск; 8 — саркомер (по Хаксли).
Комментарии к электронограмме:
См. комментарии к ЭГ №34.
На данной электронограмме представлен саркомер поперечно-полосатой миофибриллы (цифра 8). Саркомер, очевидно, находится в сокращенном состоянии, т.к. тонкие (актиновые) филаменты глубоко проникли в А-диск, поэтому I-диск и Н-полоска узкие.
37. Чувствительное инкапсулированное нервное окончание (тельце Фатера-Пачини) (рис. 201)
Пластинчатое (фатер-пачиниево) тельце. Электронная микрофотограмма.
1 — аксон (точнее, дендрит); 2 — митохондрии; 3 — щель внутренней колбы; 4 — отростки пластинчатых клеток внутренней колбы; 5 — пиноцитозные пузырьки (по В.Л.Черепнову).
Строение сердечной мышцы.
Сердечная мышца имеет клеточное строение и клеточное строение миокарда было установлено еще в 1850 году Келликером, но длительное время считалось, что миокард представляет собой сеть – сенцидий. И только электронная микроскопия подтвердила, что каждый кардиомиоцит имеет свою собственную мембрану и отделен от других кардиомиоцитов. Область контактов кардиомиоцитов – это вставочные диски. В настоящее время клетки сердечной мышцы подразделяют на клетки рабочего миокарда – кардиомиоциты рабочего миокрада предсердий и желудочков и на клетки проводящей системы сердца. Выделяют :
-P клетки – пейсмейкерные
-переходные клетки
-клетки Пуркинье
Клетки рабочего миокарда принадлежат исчерченным мышечным клеткам и кардиомиоциты имеют вытянутую форму, длин достигает 50мкм, диаметр – 10-15 мкм. Волокна состоят из миофибрилл, наименьшей рабочей структурой которых является саркомер. Последний имеет толстые — миозиновые и тонкие – актиновые ветви. На тонких нитях имеются регуляторные белки – тропанин и тропомиозин. В кардииомиоцитах имеются также продольная система L трубочек и поперечные T трубочки. Однако Т трубочки, в отличии от Т-трубочек скелетных мышц, отходят на уровне мембран Z (в скелетных — на границе диска A и I). Соседние кардиомиоциты соединяются с помощью вставочного диска- область контакта мембран. При этом структура вставочного диска неоднородная. ВО вставочном диске можно выделить область щели(10-15Нм). Вторая зона плотного контакта – десмосомы. В области десмосом наблюдается утолщение мембраны, здесь же проходят тонофибриллы(нити связывающие соседние мембраны). Десмосомы имеют протяженность 400нм. Есть плотные контакты, они получили название нексусов, при котором происходит слияние наружных слоев соседних мембран, сейчас обнаружены – конексоны – скрепление за счет специальных белко – конексинов. Нексусы – 10-13%, эта область имеет очень низкое электрическое сопротивление 1,4 Ома на кВ.см. Это обеспечивает возможность передачи электрического сигнала с одной клетки на др. и поэтому кардиомиоциты включаются одновременно в процесс возбуждения. Миокард – функциональный сенсидий.
Физиологические свойства сердечной мышцы.
Кардиомиоциты изолированы друг от друга и контактируют в области вставочных дисков, где соприкасаются мембраны соседних кардиомиоциов.
Коннесксоны- это соединение в мембране соседних клеток. Образуются эти структуры за счет белков коннексинов. Коннексон окружают 6 таких белков, внутри коннексона образуется канал, который позволяет проходит ионам, таким таким образом электрический ток распространяется от одной клетки к другой. “f область имеет сопротивление 1,4 ом на см2(низкое). Возбуждение охватывает кардиомиоциты одновременно. Они функционирую как функциональный сенсициы. Нексусы очень чувствительны к недостатку кислорода, к действию катехоламинов, к стрессовым ситуациям, к физической нагрузке. Это может вызывать нарушение проведения возбуждения в миокарде. В экспериментальных условиях нарушение плотных контактов можно получить при помещении кусочков миокарда в гипертонический раствор сахарозы. Для ритмической деятельности сердца важна проводящая система сердца – эта система состоит из комплекса мышечных клеток, образующих пучки и узлы и клетки проводящей системы отличаются от клеток рабочего миокарда – они бедны миофибриллами, богаты саркоплазмой и содержат высокое содержание гликогена. Эти особенности при световой микроскопии делают их более светлыми с малой поперечной исчерченностью и они были названы атипическими клетками.
В состав проводящей системы входят:
1. Синоатриальный узел (или узел Кейт-Фляка), расположенный в правом предсердии у места впадения верхней полой вены
2. Атриовентрикулярный узел( или узел Ашоф-Тавара), который лежит в правом предсердии на границе с желудочком — это задняя стенка правого предсердия
Эти два узла связаны внутрипредсердными трактами.
3. Предсердные тракты
— передний — с ветвью Бахмена (к левому предсердию)
— средний тракт (Венкебаха)
— задний тракт (Тореля)
4. Пучок Гисса (отходит от атриовентрикулярного узла. Проходит через фиброзную ткань и обеспечивает связь миокарда предсердия с миокардом желудочка. Проходит в межжелудочковую перегородку, где разделяется на правую и илевую ножку пучка Гисса)
5. Правая и левая ножки пучка Гисса (они идут вдоль межжелудочковой перегородки. Левая ножка имеет две ветви – переднюю и заднюю. Конечными разветвлениями будут являться волокна Пуркинье).
6. Волокна Пуркинье
В проводящей системе сердца, которая образована видоизмененными типами мышечных клеток, имеются три вида клеток : пейсмейкерные (P), переходные клетки и клетки Пуркинье.
1. P-клетки. Находятся в сино-артриальном узле, меньше в атриовентрикулярном ядре. Это самые мелкие клетки, в них мало т – фибрилл и митохондрий, т-система отсутствует, l. система развита слабо. Основной функцией этих клеток является генерация потенциала действия за счет врожденного свойства медленной диастолической деполяризации. В них происходит периодическое снижение мембранного потенциала, которое приводит их к самовозбуждению.
2. Переходные клетки осуществляют передачу возбуждения в области атривентрикуярного ядра. Они обнаруживаются между P клетками и клетками Пуркинье. Эти клетки вытянутой формы, у них отсутствует саркоплазматический ретикулум. Эти клетки облают замедленной скоростью проведения.
3. Клетки Пуркинье широкие и короткие, в них больше миофибрилл, лучше развит саркоплазматический ретикулум, T-система отсутствует.
78.30.251.74 © studopedia.ru Не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования. Есть нарушение авторского права? Напишите нам | Обратная связь.
Отключите adBlock!
и обновите страницу (F5)
очень нужно
http://studopedia.ru/19_414493_stroenie-serdechnoy-mishtsi.html
Сердечная мышца человека, ее особенности и функции
Сердце представляет собой полый орган. Его размер примерно с кулак человека. Сердечная мышца формирует стенки органа. В нем присутствует перегородка, разделяющая его на левую и правую половины. В каждой из них сеть желудочек и предсердие. Направление движения крови в органе контролируется посредством клапанов. Далее рассмотрим подробнее свойства сердечной мышцы.
Общие сведения
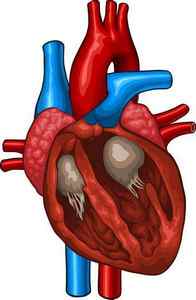
Сердечная мышца – миокард – составляет основную часть массы органа. Она состоит из трех типов ткани. В частности, выделяют: атипический миокард проводящей системы, волокна предсердия и желудочков. Размеренное и координированное сокращение сердечной мышцы обеспечивается проводящей системой.
Сердечная мышца отличается сетчатой структурой. Она формируется из волокон, переплетенных в сеть. Связи между волокнами устанавливаются за счет присутствия боковых перемычек. Таким образом, сеть представлена в виде узкопетлистого синцития. Между волокнами сердечной мышцы присутствует соединительная ткань. Она отличается рыхлой структурой. Кроме этого, волокна обвиты густой сетью капилляров.
Свойства сердечной мышцы
В структуре присутствуют вставочные диски, представленные в виде мембран, отделяющих клетки волокон друг от друга. Здесь следует отметить важные особенности сердечной мышцы. Отдельные кардиомиоциты, присутствующие в структуре в большом количестве, соединены друг с другом параллельно и последовательно. Клеточные мембраны сливаются так, что формируют щелевые контакты высокой проницаемости. Через них беспрепятственно диффундируют ионы. Таким образом, одна из особенностей миокарда состоит в наличии свободного перемещения ионов по внутриклеточной жидкости по ходу всего миокардиального волокна. Это обеспечивает беспрепятственное распределение потенциалов действия от одной клетки к другой сквозь вставочные диски. Из этого следует, что сердечная мышца – это функциональное объединение огромного количества клеток, имеющих тесную взаимосвязь друг с другом. Она настолько сильна, что при возбуждении только одной клетки провоцирует распространение потенциала на все остальные элементы.
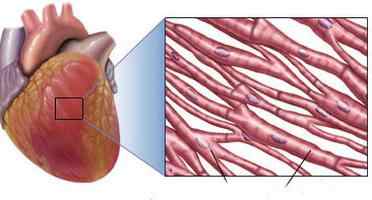
Миокардиальные синцития
В сердце их два: предсердный и желудочковый. Все отделы сердца отделены друг от друга фиброзными перегородками с отверстиями, снабженными клапанами. Непосредственно через ткань стенок возбуждение от предсердия к желудочку перейти не может. Передача осуществляется посредством специального атриовентрикулярного пучка. Его диаметр – несколько миллиметров. Состоит пучок из волокон проводящей структуры органа. Присутствие в сердце двух синцитий способствует тому, что предсердия сокращаются раньше желудочков. Это, в свою очередь, имеет важнейшее значение для обеспечения эффективной насосной деятельности органа.
Болезни миокарда
Работа сердечной мышцы может нарушаться вследствие различных патологий. В зависимости от провоцирующего фактора, выделяют специфические и идиопатические кардиомиопатии. Болезни сердца могут быть также врожденными и приобретенными. Существует еще одна классификация, в соответствии с которой различают рестриктивную, дилатационную, конгестивную и гипертрофическую кардиомиопатии. Рассмотрим их вкратце.
Гипертрофическая кардиомиопатия
На сегодняшний день специалистами выявлены мутации генов, провоцирующие данную форму патологии. Для гипертрофической кардиомиопатии характерно утолщение миокарда и изменение его структуры. На фоне патологии мышечные волокна увеличиваются в размерах, \»скручиваются\», приобретая странные формы. Первые симптомы заболевания отмечаются в детском возрасте. Основными признаками гипертрофической кардиомиопатии считаются болезненность в груди и одышка. Также наблюдается неравномерность сердечного ритма, на ЭКГ обнаруживаются изменения в сердечной мышце.
Конгестивная форма
Это достаточно распространенный тип кардиомиопатии. Как правило, заболевание возникает у мужчин. Распознать патологию можно по признакам сердечной недостаточности и нарушениям в сердечном ритме. У некоторых пациентов отмечается кровохарканье. Патологию также сопровождает боль в районе сердца.
Дилатационная кардиомиопатия
Эта форма заболевания проявляется в виде резкого расширения во всех камерах сердца и сопровождается снижением сократительной способности левого желудочка. Как правило, дилатационная кардиомиопатия возникает в сочетании с гипертонической болезнью, ИБС, стенозом в аортальном отверстии.
Рестриктивная форма
Кардиомиопатия этого типа диагностируется крайне редко. Причиной патологии является воспалительный процесс в сердечной мышце и осложнения после вмешательства на клапанах. На фоне заболевания происходит перерождение миокарда и его оболочек в соединительную ткань, отмечается замедленное наполнение желудочков. У пациента отмечается одышка, быстрая утомляемость, пороки клапанов и сердечная недостаточность. Крайне опасной рестриктивная форма считается для детей.
Как укрепить сердечную мышцу?
Существуют различные способы это сделать. Мероприятия включают в себя коррекцию режима дня и питания, упражнения. В качестве профилактики после консультации с врачом можно начать принимать ряд препаратов. Кроме этого, есть и народные методы укрепления миокарда.
Физическая активность
Она должна быть умеренной. Физическая активность должна стать неотъемлемым элементом жизни любого человека. При этом нагрузка должна быть адекватной. Не стоит перегружать сердце и истощать организм. Оптимальным вариантом считаются спортивная ходьба, плавание, езда на велосипеде. Упражнения рекомендуется проводить на свежем воздухе.
Она превосходно подходит не только для укрепления сердца, но и для оздоровления всего организма. При ходьбе задействована практически вся мускулатура человека. При этом сердце дополнительно получает умеренную нагрузку. По возможности, особенно в молодом возрасте, стоит отказаться от лифта и преодолевать высоту пешком.
Образ жизни
Укрепление сердечной мышцы невозможно без корректировки режима дня. Для улучшения деятельности миокарда необходимо отказаться от курения, дестабилизирующего давление и провоцирующего сужение просвета в сосудах. Кардиологи также не рекомендуют увлекаться баней и сауной, поскольку пребывание в парной существенно увеличивает сердечные нагрузки. Необходимо также позаботиться и о нормальном сне. Спать следует ложиться вовремя и отдыхать достаточное количество часов.
Одним из важнейших мероприятий в вопросе укрепления миокарда считается рациональное питание. Следует ограничить количество соленой и жирной пищи. В продуктах должны присутствовать:
- Магний (бобовые, арбузы, орехи, гречка).
- Калий (какао, изюм, виноград, абрикосы, кабачки).
- Витамины Р и С (клубника, черная смородина, перец (сладкий), яблоки, апельсины).
- Йод (капуста, творог, свекла, морепродукты).
Негативное воздействие на деятельность миокарда оказывает холестерин в высоких концентрациях.

Психоэмоциональное состояние
Укрепление сердечной мышцы может осложняться различными неразрешенными проблемами личного либо рабочего характера. Они могут спровоцировать перепады давления и нарушения ритма. Следует по возможности избегать стрессовых ситуаций.
Существует несколько средств, способствующих укреплению миокарда. К ним, в частности, относят такие препараты, как:
- \»Рибоксин\». Его действие направлено на стабилизацию ритма, усиление питания мышцы и коронарных сосудов.
- \»Аспаркам\». Этот препарат представляет собой магниево-калиевый комплекс. Благодаря приему средства нормализуется электролитный обмен, устраняются признаки аритмии.
- Родиола розовая. Это средство улучшает сократительную функцию миокарда. При приеме данного препарата следует соблюдать осторожность, поскольку он обладает способностью к возбуждению нервной системы.
http://www.syl.ru/article/169142/new_serdechnaya-myishtsa-cheloveka-ee-osobennosti-i-funktsii
МЫШЕЧНОЙ ТКАНИ. Сердечная мышечная тканьобразована клетками — кардиомиоцитами(сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими;
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ СЕРДЕЧНОЙ
Сердечная мышечная тканьобразована клетками — кардиомиоцитами(сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон (рис. 7.12).
Кардиомиоциты — цилиндрические или ветвящиеся клетки, более крупные в желудочках, где их длина составляет 100-150 мкм, а диаметр — 10-20 мкм. В предсердиях они обычно имеют неправильную форму и меньшие размеры (длина — 40-70 мкм, диаметр — 5-6 мкм). Кардиомиоциты содержат одно или два ядра и саркоплазму, покрыты сарколеммой, которая снаружи окружена базальной мембраной.
Саркоплазма кардиомиоцитов содержит органеллы и включения, которые образуют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппаратсильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых), которых он занимает до 50-70% объема клетки. Фагоструктура сходна с таковой поперечно-полосатой мышечной ткани. Механизм мышечного сокращения этих видов мышечной ткани также сходны.
Рис. 7.13. Саркотубулярная система кардиомиоцита. Саркотубулярная система представлена саркоплазматической сетью (СПС), не образующей терминальных цистерн, и широкими Т-трубочками (Т-ТР).
Аппарат передачи возбуждения (саркотубулярная система)в кардиомиоцитах в целом сходен с таковым в скелетных мышечных волоконах, однако он обладает рядом особенностей (рис. 7.13).
Саркоплазматическая сеть развита слабее, чем в скелетном мышечном волокне, менее активно накапливает Са 2+ , не образует терминальных цистерн. Во время расслабления она выделяет ионы Са 2+ в саркоплазму с низкой скоростью, что обеспечивает автоматизм кардиомиоцитов (рис. 7.14).
Рис. 7.14. На электронной микрофотографии изображен: А (х5.000) – участок миокарда состоящий из 6 миоцитов (С1-С6). Эти клетки соединяются между собой вставочным диском D. В поле зрения в миоцитах нет ядер. Между миоцитами – рыхлая соединительная ткань, в которой много капилляров (Сар), фибробластов (F); Б (х38.000) ультраструктура саркомера миокарда схожа с таковой поперечно-полосатой мышечной тканью. Обратить внимание на строение А и J дисков СПР (SR), тубулярной системы (Т), расположение митохондрий. В миоците много липидных капель (L) и гранул гликогена (G).
Поперечные (Т-) трубочки — широкие, содержат компоненты базальной мембраны, вместе с элементами саркоплазматической сети образуют диады (включают одну
Т-трубочку и одну цистерну сети), которые располагаются в области Z-линий (рис. 7.14).
Опорный аппарат кардиомиоцитовпредставлен элементами цитоскелета, обеспечивающими упорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также базальной мембраной и сарколеммой. Его структурная и биохимическая организация сходна с таковой в волокнах скелетной мышечной ткани. В кардиомиоцитах его особенностью служит то, что элементы цитоскелета связаны с особыми межклеточными соединениями – вставочными дисками.
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих функциональное волокно сердечной мышечной ткани (рис. 7.15).
Энергетический аппарат кардиомиоцитовпредставлен митохондриями и включениями, расщепление которых обеспечивает получение энергии. Митохондрии кардиомиоцитов обладают также свойством накапливать кальций в высоких концентрациях.
Рис. 7.15. Ультраструктурная организация области вставочного диска кардиомиоцитов. В поперечных участках вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом (Д) актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска в участке полоски слипания (ПС). На сарколемме продольных участков вставочного диска располагаются щелевые соединения (ЩС). БМ – базальная мембрана, СЛ – сарколемма, МТХ – митохондрия. Обозначения компонентов саркомера СМ) – те же, что на рис. 4.
Синтетический аппарат кардиомиоцитов Синтетический аппарат включает свободные рибосомы и полисомы, цистерны грЭПС. и элементы комплекса Гольджи, которые располагаются в саркоплазме преимущественно у полюсов ядра.
Лизосомальный аппарат (аппарат внутриклеточного переваривания) кардиомиоцитовхорошо развит, что отражает высокую скорость обновления их структурных компонентов. Он включает эндосомы, лизосомы и остаточные тельца. Собственно лизосомы располагаются у полюсов ядра и занимают до 10% объема саркоплазмы. Остаточные тельца в виде липофусциновых гранул очень многочисленны (при старении могут составлять до 20% сухой массы миокарда).
http://studopedia.su/15_161893_mishechnoy-tkani.html
В области вставочных дисков сердечной мышцы имеются
3) зоны прикрепления миофибрилл
4) интердигитации (пальцевидные впячивания контактирующих
Гладкие мышечные клетки способны синтезировать
IV. ДОПОЛНИТЕ
4. Камбиальные элементы скелетной мышечной ткани______________.
V. ГРАФИЧЕСКИЕ ЗАДАНИЯ (укажите положение на рисунках)
Укажите на рисунке положение составных частей саркомера

ТЕМА «МЫШЕЧНЫЕ ТКАНИ»
Вариант №2
I. ВЫБЕРИТЕ ОДИН ПРАВИЛЬНЫЙ ОТВЕТ
а) \»M\» линиями (мезофрагмами)
б) \»Z\» линиями (телофрагмами)
а) водитель ритма (пейсмекер)
б) кардиомиоцит волокон Пуркинье
в) предсердный кардиомиоцит
г) желудочковый кардиомиоцит
д) кардиомиоцит пучка Хиса
а) под сарколеммой
б) между базальной мембраной и плазмолеммой мышечного волокна
в) между базальной мембраной и эндомизием
II. ПОДБЕРИТЕ СООТВЕТСТВУЮЩИЕ ОТВЕТЫ (ПАРЫ)
ВЫБЕРИТЕ, ПРИ КАКОМ УСЛОВИИ ВЕРНО ДАННОЕ УТВЕРЖДЕНИЕ
2) замыкательными пластинками
1) оксифилией цитоплазмы клеток
2) наличием одного — двух ядер в центре клетки
3) наличием вставочных дисков
4) наличием большого количества соединительной ткани между клетками
4) мышечное волокно
V. ГРАФИЧЕСКИЕ ЗАДАНИЯ (укажите положение на рисунках)
Рабочие кардиомиоциты
Кардиомиоциты — морфо-функциональные единицы сердечной мышечной ткани, имеют цилиндрическую ветвящуюся форму диаметром около 15 мкм (рис. 7-22). При помощи межклеточных контактов (вставочные диски) рабочие кардиомиоциты объединены в так называемые сердечные мышечные волокна — функциональный синцитий (рис. 7-21А) — совокупность кардиомиоцитов в пределах каждой камеры сердца. Клетки содержат центрально расположенные, вытянутые вдоль оси одно или два ядра, миофибриллы и ассоциированные с ними цистерны саркоплазматического ретикулума (депо Ca 2+ ). Многочисленные митохондрии залегают параллельными рядами между миофибриллами. Их более плотные скопления наблюдают на уровне I-дисков и ядер. Гранулы гликогена сконцентрированы на обоих полюсах ядра. Митохондрии и запасы гликогена поддерживают окислительный метаболизм. Т-трубочки в кардиомиоцитах — в отличие от скелетных мышечных волокон — проходят на уровне Z-линий. В связи с этим Т-трубочка контактирует только с одной терминальной цистерной. В результате вместо триад скелетного мышечного волокна формируются диады.
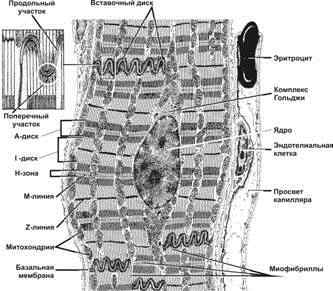
Рис.7—22.Рабочий кардиомиоцит — удлинённой формы клетка. Ядро расположено центрально, вблизи ядра находятся комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Вставочные диски (на врезке) служат для скрепления кардиомиоцитов и синхронизации их сокращения. [17]
Сократительный аппарат.Организация миофибрилл и саркомеров в кардиомиоцитах такая же, что и в скелетном мышечном волокне. Одинаков и механизм взаимодействия тонких и толстых нитей при сокращении.
? Сердечные миозины. Существует несколько генов для лёгких и тяжёлых цепей сердечного миозина, различающихся по активности АТФазы. В зрелом миокарде преимущественно синтезируется тяжёлая ??цепь сердечного миозина с низкой АТФазной активностью, в отличие от ??цепи, обладающей высокой АТФазной активностью.
Гипертрофическая кардиомиопатия — гетерогенная группа наследственных заболеваний. Мутации генов тяжёлой ??цепи сердечного миозина (40%), С–белка (42%), TnT, ??тропомиозина сопровождаются гипертрофией кардиомиоцитов и дезорганизацией сердечных мышечных волокон.
? Тропонины. Изоформа тропонина С сходна с TnC1, экспрессирующегося в медленных скелетных мышечных волокнах. TnI характеризуется высокой специфичностью для сердечной мышцы.
Саркоплазматическая сеть. Выход Ca 2+ из саркоплазматического ретикулума регулируется через рецепторы рианодина (RyR2). Изменения мембранного потенциала открывают потенциалозависимые Ca 2+ -каналы, в кардиомиоцитах незначительно повышается концентрация Ca 2+ . Этот Ca 2+ активирует рецепторы рианодина, и Ca 2+ из депо выходит в цитозоль (кальций-индуцированная мобилизация Ca 2+ ). В момент расслабления кардиомиоцитов Ca 2+ -АТФаза SERCA2a закачивает Ca 2+ в саркоплазматический ретикулум, где ионы Са 2+ связывает сердечная изоформа белка кальсеквестрина. Фосфоламбан регулирует активность SERCA2a. Нефосфорилированная форма фосфоламбана блокирует SERCA2a. Норадреналин через цАМФ-зависимую протеин киназу А путём фосфорилирования фосфоламбана снимает ингибирующий эффект фосфоламбана на Ca 2+ -АТФазу, облегчая таким образом процесс расслабления мышечного волокна и увеличивая эффективность сокращения. Для предотвращения избыточного депонирования Ca 2+ те катионы, которые проникли в клетку через плазмолемму, должны быть выведены во внеклеточное пространство.Натрий-кальциевый анионообменник осуществляет обмен внутриклеточного Ca 2+ на внеклеточный Na + .
? Мутация гена фосфоламбана является одной из причин дилатационной кардиомиопатии, характеризующейся диффузным поражением миокарда с дилатацией (расширением) полостей сердца и резким снижением сократительной функции сердца.
? Йодсодержащие гормоны Т3 и Т4 активируют гены, кодирующие Са 2+ -АТФазу, Na + ,K + -АТФазу, ?1-адренорецептор, транспортёр глюкозы, предсердный натриуретический пептид. При повышенном содержании в крови Т3 и Т4 в кардиомиоцитах подавляется синтез тяжёлой ?-цепи и увеличивается экспрессия гена, кодирующего тяжёлую ?-цепь сердечного миозина.
Вставочные диски. На концах контактирующих кардиомиоцитов имеются интердигитации (пальцевидные выпячивания и углубления). Вырост одной клетки плотно входит в углубление другой. На конце такого выступа (поперечный участок вставочного диска) сконцентрированы контакты двух типов: десмосомы и промежуточные. На боковой поверхности выступа (продольный участок вставочного диска) имеется множество щелевых контактов (nexus, нексус), передающих возбуждение от кардиомиоцита к кардиомиоциту.
? Десмосомы обеспечивают механическое сцепление, препятствующее расхождению кардиомиоцитов при сокращении.
? Промежуточные контакты необходимы для прикрепления тонких актиновых нитей ближайшего сaркомера к сарколемме кардиомиоцита.
? Щелевые контакты (nexus) — межклеточные ионные каналы — образуют электрические синапсы, проводящие возбуждение от кардиомиоцита к кардиомиоциту. Это обстоятельство — наряду с проводящей системой сердца — позволяет синхронизировать одновременное сокращение множества кардиомиоцитов в составе функционального синцития.
Синхронизация сокращения кардиомиоцитов. Распространение волны деполяризации по сердечным мышечным волокнам обеспечивают электрические синапсы между смежными кардиомиоцитами. Внутриклеточный и внеклеточный потоки ионов, обеспечивающие генерацию потенциала действия и синхронизацию сокращения сердечной мышцы, лежат в основе электромиографического исследования миокарда (рис. 7-23).
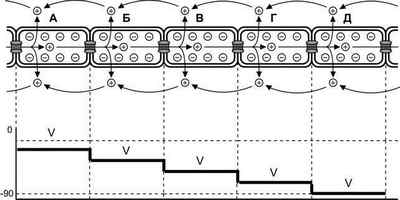
Рис. 7-23. Проведение возбуждения между кардиомиоцитами.Положительно заряженные ионы из кардиомиоцита А, в котором возник потенциал действия, через электрические синапсы переходят в смежные клетки (Б, В, Г, Д), вызывая в них деполяризацию (V). Степень деполяризации кардиомиоцитов зависит от расстояния между клетками и приближающейся волной возбуждения. В кардиомиоците (Б) деполяризация активирует потенциалозависимые Na + — и Ca 2+ -каналы. Поступившие в цитозоль внеклеточные Na + и Ca 2+ приближают деполяризацию мембраны к критическому порогу возникновения потенциала действия и увеличивают внутриклеточный поток положительно заряженных ионов, распространяющийся далее по клеткам (В, Г, Д) впереди волны возбуждения. При этом в результате деполяризации ассоциированные с мембраной положительно заряженные ионы выходят из кардиомиоцитов во внеклеточное пространство и перемещаются в обратном внутриклеточному току направлении. [114]
Предсердные и желудочковые кардиомиоциты. Предсердные и желудочковые кардиомиоциты относятся к разным популяциям рабочих кардиомиоцитов. Предсердные кардиомиоциты относительно мелкие, 10 мкм в диаметре и длиной 20 мкм. В них слабее развита система Т-трубочек, но в зоне вставочных дисков значительно больше щелевых контактов. Желудочковые кардиомиоциты крупнее (25 мкм в диаметре и до 140 мкм в длину), они имеют хорошо развитую систему Т-трубочек. В состав сократительного аппарата миоцитов предсердий и желудочков входят разные изоформы миозина, актина и других контрактильных белков.
Секреторные кардиомиоциты. В части кардиомиоцитов предсердий (особенно правого) у полюсов ядер располагаются хорошо выраженный комплекс Гольджи и секреторные гранулы, содержащие атриопептин — гормон, регулирующий АД. При повышении АД стенка предсердия сильно растягивается, что стимулирует предсердные кардиомиоциты к синтезу и секреции атриопептина, вызывающего снижение АД.
Дата добавления: 2015-07-07 ; просмотров: 2626 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
http://helpiks.org/4-5733.html
Вставочные диски в сердечной мышце
Здравствуйте читатели моего проекта \»Биология для студентов\»! Подготовка к экзаменам, зачетам и госэкзаменам, а также рефераты и презентации занимают много времени, если готовится по учебникам. Есть три способа подготовки к экзамену: по учебнику, по лекциям и поиск в интернете. Готовиться по учебнику очень долго. Что касается лекций, не у всех есть хорошие лекции, так как не все преподаватели их нормально читают, и кроме того не все успевают их записывать. И остается третий вариант искать ответы на вопросы в интернете. Не для кого не секрет, что в настоящее время большинство студентов предпочитают именно этот вариант.

За пять лет учебы на факультете биотехнологии и биологии подготовка к сессии у меня занимала много времени. В Рунете не так много биологических сайтов. Конспекты по экономике, истории, социологии, политологии, математике найти очень просто. А ответы на вопросы по ботанике, зоологии, генетики, биофизике, биохимии гораздо сложнее. Наверное, потому что биология не самая распространенная специальность. К тому же биологические предметы не являются общеобразовательными в отличие, например, от экономики и истории, которые изучаются практически на любых специальностях. В Рунете я не нашел ни одного сайта на которым был бы представлен необходимый контент для подготовки к экзаменам, зачетам и госэкзаменам по биологическим дисциплинам. И я решил создать его.
Также я хотел бы вас попросить рассказать об этом сайте своим однокурсникам, друзьям и знакомым, которые являются студентами биологических специальностей. Это поможет развитию данного проекта.
Под ред. Э.Г. Улумбекова, Ю.А. Челышева, Гистология (введение в патологию), 1997
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Поперечнополосатая мышечная ткань сердечного типа входит в состав мышечной стенки сердца (миокард). Основной гистологический элемент — кардиомиоцит. Кардиомиоциты присутствуют также в проксимальной части аорты и верхней полой вены.
А. Кардиомиогенез. Миобласты происходят из клеток спланхнической мезодермы, окружающей эндокардиальную трубку (глава 10 Б I). После ряда митотических делений G,-mho6- ласты начинают синтез сократительных и вспомогательных белков и через стадию G0- миобластов дифференцируются в кардиомиоциты, приобретая вытянутую форму; в саркоплазме начинается сборка миофибрилл. В отличие от поперечнополосатой мышечной ткани скелетного типа, в кардиомиогенезе не происходит обособления камбиального резерва, а все кардиомиоциты необратимо находятся в фазе G0 клеточного цикла. Специфический фактор транскрипции (ген CATFl/SMBP2, 600502, Ilql3.2-ql3.4) экспрессируется только в развивающемся и сформировавшемся миокарде.
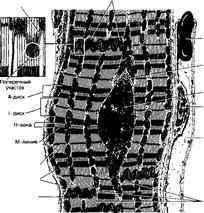
Б. Кардиомиоциты расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры бассейна венечных сосудов и терминальные ветвления двигательных аксонов нервных клеток вегетативного отдела нервной системы. Каждый миоцит имеет сарколемму (базальная мембрана + плазмолемма). Различают рабочие, атипичные и секреторные кардиомиоциты.
а. Сократительный аппарат. Организация миофибрилл и саркомеров в кардиомио- цитах такая же, как и в скелетном мышечном волокне (см. I Б I, 2). Одинаков и механизм взаимодействия тонких и толстых нитей при сокращении (см. I Г 5, 6, 7).
б. Саркоплазматическая сеть. Выброс Ca2+ из саркоплазматического ретикулума регулируется через рецепторы рианодина (см. также главу 2 III А 3 б (3) (а)). Изменения мембранного потенциала открывают потенциалзависимые Са2+-каналы, в кар- диомиоцитах незначительно повышается концентрация Ca2+. Этот Ca2+ активирует рецепторы рианодина, и Ca2* выходит в цитозоль (кальций-индуцированная мобилизация Ca2+).
в. Т-трубочки в кардиомиоцитах, в отличие от скелетных мышечных волокон, проходят на уровне Z-линий. В связи с этим Т-трубочка контактирует только с одной терминальной цистерной. В результате вместо триад скелетного мышечного волокна формируются диады.
г. Митохондрии расположены параллельными рядами между миофибриллами. Их более плотные скопления наблюдают на уровне I-дисков и ядер.
Комплекс Г ольджи
Ядро
Эндотелиальная
клетка
Рис. 7-11. Рабочий кардиомиоцит — удлинённой формы клетка. Ядро расположено центрально, вблизи ядра находятся комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Вставочные диски (на врезке) служат для скрепления кардиомиоцитов и синхронизации их сокращения [из Hees H, Sinowatz F (1992) и Kopf-MaierP, Merker H-J <1989))
д. Вставочные диски. На концах контактирующих кардиомиоцитов имеются интердигитации (пальцевидные выпячивания и углубления). Вырост одной клетки плотно входит в углубление другой. На конце такого выступа (поперечный участок вставочного диска) сконцентрированы контакты двух типов: десмосомы и промежуточные. На боковой поверхности выступа (продольный участок вставочного диска) имеется множество щелевых контактов (nexus, нексус).
е. Предсердные и желудочковые миоциты — разные популяции рабочих кардиомиоцитов. В предсердных кардиомиоцитах слабее развита система Т-трубочек, но в зоне вставочных дисков значительно больше щелевых контактов. Желудочковые кардиомиоциты крупнее, они имеют хорошо развитую систему Т-трубочек. В состав сократительного аппарата миоцитов предсердий и желудочков входят разные изоформы миозина, актина и других контрактильных белков.
а. Водители ритма (пейсмейкерные клетки, пейсмейкеры; рис. 7-12) — совокупность специализированных кардиомиоцитов в виде тонких волокон, окружённых рыхлой соединительной тканью. По сравнению с рабочими кардиомиоцитами они имеют меньшие размеры. В саркоплазме содержится сравнительно мало гликогена и небольшое количество миофибрилл, лежащих в основном по периферии клеток. Эти клетки имеют богатую васкуляризацию и двигательную вегетативную иннервацию. Так, в синусно- предсердном узле доля соединительнотканных элементов (включая кровеносные капилляры) в 1,5-3 раза, а нервных элементов (нейроны и двигательные нервные окончания) в 2,5-5 раз выше, чем в рабочем миокарде правого предсердия. Главное свойство водителей ритма — спонтанная деполяризация плазматической мембраны. При достижении критического значения возникает потенциал действия, распространяющийся по волокнам проводящей системы сердца и достигающий рабочих кардиомиоцитов. Главный водитель ритма — клетки синусно-предсердного узла — генерирует ритм 60-90 импульсов в минуту. Нормально активность других водителей ритма подавлена.
б. Проводящие кардиомиоциты — специализированные клетки, выполняющие функцию проведения возбуждения от водителей ритма. Эти клетки образуют длинные волокна.
В. Иннервация. На деятельность сердца — сложной авторегуляторной и регулируемой системы — оказывает влияние множество факторов, в т.ч. двигательная вегетативная
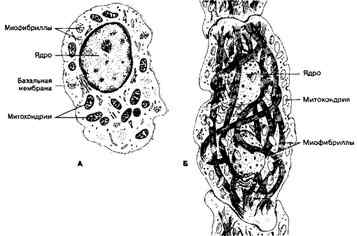
Рис. 7-12. Атипичные кардиомиоциты. А — водитель ритма синусно-предсердного узла;
Б — проводящий кардиомиоцит пучка Гйса [из Hees Н, Sinowatz F, 1992]
иннервация — парасимпатическая и симпатическая. Парасимпатическая иннервация осуществляется терминальными варикозными окончаниями аксонов блуждающего нерва, а симпатическая — окончаниями аксонов адренергических нейронов шейного верхнего, шейного среднего и звёздчатого (шейно-грудного) ганглиев. В контексте представления о сердце как о сложной авторегуляторной системе чувствительная иннервация сердца (как вегетативная, так и соматическая) должна рассматриваться как часть системы регуляции
кровотока.
адренергические рецепторы плазмолеммы разных клеток сердца (кардиомиоциты рабочие и особенно атипические, внутрисердечные нейроны собственного нервного аппарата). Существует множество фармакологических препаратов, оказывающих непосредственное действие на названные рецепторы. Так, норадреналин, адреналин и другие адренергические препараты в зависимости от эффекта на а- и p-адренорецепторы подразделяют на активирующие (адреномиметики) и блокирующие (адреноблока- торы) агенты. м-Холинорецепторы также имеют аналогичные классы препаратов (холиномиметики и холиноблокаторы).
а. Активация симпатических нервов увеличивает частоту спонтанной деполяризации мембран водителей ритма, облегчает проведение импульса в волокнах Пуркинье и увеличивает частоту и силу сокращения типичных кардиомиоцитов.
б. Парасимпатические влияния, наоборот, уменьшают частоту генерации импульсов пейсмейкерами, снижают скорость проведения импульса в волокнах Пуркинье и уменьшают частоту сокращения рабочих кардиомиоцитов.
а. Спинальная. Периферические отростки чувствительных нейронов спинномозговых узлов образуют свободные и инкапсулированные нервные окончания.
б. Специализированные сенсорные структуры сердечно-сосудистой системы рассмотрены в главе 10.
Г. Регенерация. При ишемической болезни сердца (ИБС), атеросклерозе коронарных сосудов, сердечной недостаточности разной этиологии (в т.ч. при артериальной гипертензии, инфаркте миокарда) наблюдаются патологические изменения кардиомиоцитов, включая их гибель.
а. Причины сердечной недостаточности — снижение сократительной способности, увеличение посленагрузки, изменения преднагрузки.
Снижение сократительной способности
(а) Инфаркт миокарда — некроз участка сердечной мышцы с потерей его способности к сокращению. Замещение поражённой части стенки желудочков соединительной тканью приводит к снижению функциональных свойств миокарда. При поражении значительной части миокарда развивается сердечная недостаточность.
(б) Врождённые и приобретённые пороки сердца приводят к перегрузке полостей сердца давлением или объёмом с развитием сердечной недостаточности.
(в) Артериальная гипертензия. Многие больные гипертонической болезнью или симптоматическими гипертензиями страдают недостаточностью кровообращения. Снижение сократительной способности миокарда характерно для стойкой тяжёлой гипертензии, быстро приводящей к развитию сердечной недостаточности.
(г) Кардиомиопатии токсические (алкоголь, кобальт, катехоламины, доксору- бицин), инфекционные, при т.н. коллагеновых болезнях, рестриктивные (ами- лоидоз и саркоидоз, идиопатические).
б. Компенсаторные механизмы при сердечной недостаточности. Феномены, вытекающие из закона Франка-Старлинга, в т.ч. гипертрофия миокарда, дилатация левого желудочка, периферическая вазоконстрикция вследствие выброса катехоламинов, активация системы ренин-ангиотензин-[альдостерон] и вазопрессина, перепрограммирование синтеза миозинов в кардиомиоцитах, увеличение секреции атриопептина, — компенсаторные механизмы, поддерживающие положительный инотропный эффект. Однако рано или поздно миокард теряет способность обеспечивать нормальный сердечный выброс.